

-
Global/EN
- Global
- North America
- Latin America
Exogenous phytases have been used commercially since the early 1990s as a successful tool for reducing the environmental impact of industrial livestock production and improving poultry and swine profitability (Selle & Ravindran, 2007). These cost-saving and sustainability benefits derive primarily from phytase’s ability to liberate phosphorus from phytate. The breakdown of this poorly digestible compound improves animals’ phytate-phosphorus retention and reduces the need to use inorganic phosphorus sources in the diet.
However, the hydrolysis of phytate also delivers several additional physiological effects in animals, which extend beyond phosphorus alone. These additional benefits include the retention of amino acids, trace minerals, calcium and energy. Phytase also offers performance enhancements which extend beyond expectations associated simply with nutrient release values (Cowieson et al., 2011). The exact cause of these enhanced benefits, however, is currently not fully understood. New research conducted by DSM has revealed important new insights into the ‘extra-phosphoric’ effects of phytase, particularly on the role of myo-inositol.
Phytate is the main phosphorus store of many plants and a dietary anti-nutrient. It reduces the solubility of protein and various cations through electrostatic mechanisms, impeding digestion and increasing endogenous nutrient loss (Cowieson & Ravindran, 2007). These negative effects can be substantial, although the exact magnitude depends on a variety of factors, including phytate concentration, protein source and cation/anion balance (Cowieson et al., 2011; Bye et al., 2013).
It is well known that phytase improves the digestibility of phosphorus and reduces the antinutritive effects of phytate. However, numerous studies have identified that its effects on live performance go well beyond what could be logically explained by these two activities alone. This is particularly true for so-called phytase ‘super-dosing’ (Cowieson et al., 2011).
Phytase has traditionally been used to reduce diet costs by replacing inorganic phosphate sources. It has also occasionally been used to substitute energy sources such as animal or vegetable fat, as well as to allow the reduction of limestone, synthetic amino acids and salts in diets. The effectiveness of such displacements can be linked to a nutrient release matrix for a particular product at a defined inclusion concentration. The value derived will depend simply on the ‘shadow’ prices of the various replaced nutrients and ingredients – a measurement which can be readily calculated.
Like all enzymes, the effects of phytase dosing follow a log-linear response curve. Doubling a standard dose will result in an increase in efficacy of only around 30 per cent and doubling a second time will provide an additional increment of just 18 per cent (Selle & Ravindran, 2007). This log-linear response therefore makes it difficult to justify unconventionally high doses. In theory, the return-on-investment should decline with each unit of activity added.
However, as the value created by phytase may extend beyond the benefits derived from nutrient release alone, such approaches do not take into account the full impact of phytase supplementation. Research attention has therefore begun to shift away from phosphorus release and anti-nutritive reductions, to focus more prominently on the role of myo-inositol.
Myo-inositol is a cyclical sugar alcohol with a formula similar to glucose, which forms the core of a phytate molecule. The role of myo-inositol in nutrition is not clear and is an active area for research, particularly in human dietetics. However, myo-inositol has been found to have similar properties and metabolic functions to insulin, stimulating the translocation of GLUT4 (the primary insulin-sensitive glucose transporter in mammals) to the plasma membrane. This suggests that it may regulate glucose transport, gluconeogenesis and protein deposition in mammals (Dang et al., 2010; Yamashita et al., 2013).
However, until recently the effects of myo-inositol in poultry have been thought to differ from that of mammals, as avian species generally lack GLUT4. Recent research has challenged this conclusion, however, finding that many species of bird, including poultry, are actually insulin responsive (Tokushima et al., 2005; Sweazea & Braun, 2006). Indeed, orally administered myo-inositol has been shown to improve the performance of broiler chicks (Cowieson et al., 2013; Zyla et al., 2013).
To further clarify the exact role of myo-inositol in the phytase response of poultry, two studies were recently undertaken by DSM to examine the effect of RONOZYME® HiPhos GT (a commercially available phytase) on plasma myo-inositol concentrations (Cowieson et al., 2014).
In both experiments, a group of Ross broiler chicks was raised for a period of three to four weeks post-hatch on corn/soy-based diets containing insufficient available phosphorus and calcium (Negative Control (NC)). An additional control group was fed a diet containing sufficient phosphorus and calcium (Positive Control (NC)). Both diets were supplemented with RONOZYME® HiPhos, from 1000-3000 FYT/kg.
In addition to standard performance metrics (i.e. bone characteristics and mineral retention), plasma myo-inositol concentrations were measured by mass spectrometry using an UPLC-TQD system (Water, MA 01757, Milford, USA). These measurements were recorded based on the method described by Leung et al. (2011).
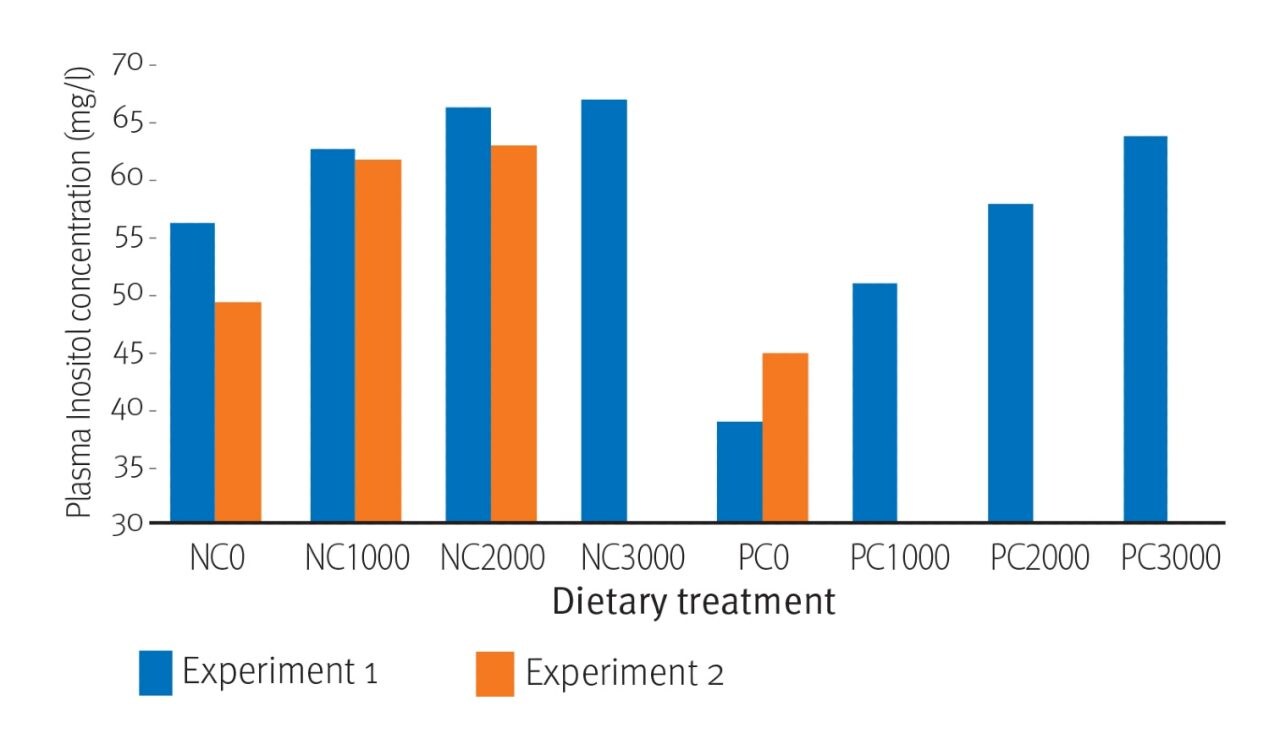 Fig. 1. Effect of RONOZYME® HiPhos GT (expressed as FYT/kg) on plasma myo-inositol concentrations in broiler chicks fed diets containing insufficient (NC) and sufficient (PC) available P and Ca levels.
Fig. 1. Effect of RONOZYME® HiPhos GT (expressed as FYT/kg) on plasma myo-inositol concentrations in broiler chicks fed diets containing insufficient (NC) and sufficient (PC) available P and Ca levels.
In general, the performance, bone strength and mineral retention effects of phytase were as expected (Cowieson et al., 2014). Phytase improved (P<0.01) weight gain, feed conversion ratio, tibia ash and the retention of phosphorus, calcium and sodium. Additionally, phytase addition increased (P<0.001) plasma myo-inositol concentrations from around 50mg/l to almost 70mg/l (Fig.1). These effects were reflected in improved bodyweight gain (Fig. 2).
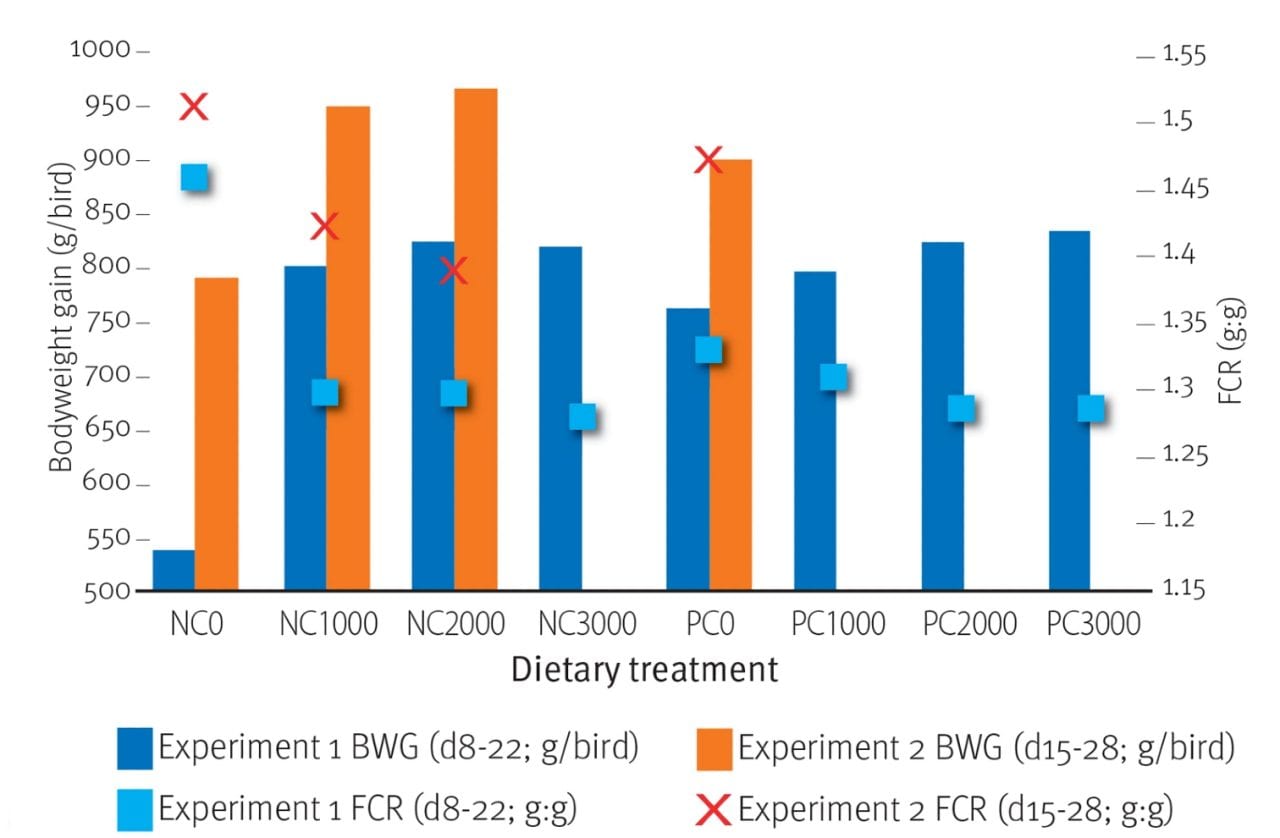 Fig. 2. Effect of RONOZYME® HiPhos GT (expressed as FYT/kg) on bodyweight gain and feed conversion ratio in broiler chicks fed diets containing insufficient (NC) and sufficient (PC) available P and Ca levels
Fig. 2. Effect of RONOZYME® HiPhos GT (expressed as FYT/kg) on bodyweight gain and feed conversion ratio in broiler chicks fed diets containing insufficient (NC) and sufficient (PC) available P and Ca levels
The impact of phytase on broiler performance, phosphorus and calcium digestibility and bone ash has been extensively reported over the past two decades (Selle et al., 2009). Results from the current studies support previous research in proving that phytase improves the retention of phosphorus and calcium, as well as bone metrics. The studies also confirm previous findings that phytase supplementation enhances the performance of birds fed a diet with a sub-optimal supply of available phosphorus and calcium.
However, the results from the control group revealed that adding phytase to a diet containing sufficient phosphorus and calcium promotes improved bodyweight gain, suggesting extraphosphoric effects may be responsible. It is possible that phytase addition, to both the PC and NC diets, resulted in increased amino acid digestibility which may be associated with reduced endogenous amino acid flow (Cowieson & Ravindran, 2007). However, the increased concentration of myo-inositol in the plasma of birds fed diets containing phytase suggest that some of the 'extraphosphoric' effects may be mediated via mechanisms involving myo-inositol.
The effect of phytase addition on plasma myo-inositol concentrations was clearly demonstrated in both studies. Indeed, concentrations increased from 39mg/l in PC0 to 67mg/l in NC3000 (71%; P<0.001) in the first experiment and from 45mg/l in PC0 to 63mg/l in NC2000 (40%; P<0.001) in the second (Fig.1). Interestingly, in the first experiment, plasma myo-inositol concentration was increased (43%; P<0.01) from 39mg/l to 56mg/l by the removal of calcium and available phosphorus from the diet (found by comparing the NC and PC groups). This effect may be associated with an increase in phytate-phosphorus digestibility in the NC group compared with the PC group, as low calcium and phosphorus concentrations often result in increased levels of phosphorus digestibility.
Previous studies have shown that low calcium concentrations assist phytate-phosphorus digestibility via increased and prolonged solubility of phytate and presumed cooperativity with mucosal phosphatases in the small intestine. (Tamim et al., 2004; Wilkinson et al., 2013). DSM’s studies confirm that the effect of dietary calcium and available phosphorus on myo-inositol is of importance. The results also show that diets with a high basal concentration of calcium may have depressed myo-inositol concentrations.
Existing DSM data suggests that baseline plasma myo-inositol concentrations in broilers are around 30mg/l and in pigs around 5-10mg/l. These data also indicate that these values are lowest in animals fed high calcium rations (Guggenbuhl et al., 2013; Cowieson et al., 2014). Compromised performance and nutrient digestibility in high calcium diets (Selle et al., 2010) may therefore be linked to depressed phytate solubility and lower plasma myo-inositol concentrations. The increase in plasma myo-inositol associated with phytase addition is a central discovery in DSM’s new studies and may explain why the effects of phytase often extend beyond those expected by the nutrient release values.
As mentioned earlier, myo-inositol mimics the role of insulin. It has the same effect on GLUT4 when blood glucose concentrations rise, controlling circulating glucose concentrations. However, a complication in understanding how this improves performance for poultry is that avian species do not possess GLUT4, meaning that this mimicking effect should theoretically be restricted to pigs and other mammals.
Chickens are, however, insulin-sensitive and it is likely that there are alternative glucose transporters in birds that are involved in these effects (Tokushima et al., 2005; Sweazea & Braun, 2006). As phytase increases myo-inositol in the plasma of chicks, it may be that this has an insulin-like effect, possibly via stimulation of pathways downstream of insulin and IGF-1 such as phosphatidylinositol-3-kinase, Akt and mTOR (pathways responsible for protein accretion). This mechanism may explain why phytase addition – particularly at high inclusion concentrations – has positive benefits on body weight gain and feed conversion ratio that extend beyond the sum of the effects on nutrient digestibility.
DSM’s research has provided revealing new insights into the effectiveness of microbial phytase in improving calcium and phosphorus digestibility and enhancing performance, when birds are fed diets insufficient in calcium and available phosphorus. The new data suggests that the extraphosphoric effects of phytase may extend from improved amino acid and trace mineral digestibility to substantial increases in plasma myo-inositol concentration.
Although further research is required to assess the importance of myo-inositol for chicks and its role in mimicking insulin, a convergent body of scientific evidence has confirmed the important role of myo-inositol in promoting growth. While phytase has historically been considered an effective tool for improving phosphorus retention and reducing the anti-nutritive effects of phytate, a third mechanism has now emerged that may be central to the beneficial effects observed.
04 September 2018

Aaron is a Corporate Science Fellow. He has a PhD in Animal Nutrition obtained from Aberdeen University, UK. Aaron has worked in commercial and academic innovation leadership roles since 2001, most recently as a Professor at the University of Sydney in Australia, joining the DSM innovation team in 2013. Aaron has published more than 200 peer-reviewed papers and book chapters and in addition to his role in DSM, is retained by Purdue University as an Adjunct Professor. In 2016 Aaron was awarded the Poultry Nutrition Research Award by the American Feed Industry Association and in 2019 the Life Mentor Award by the Poultry Science Association.

Fidelis is the Vice President/Business Director for EMEA within Animal Nutrition and Health at DSM. He holds a PhD in Animal Nutrition (University of Bonn) and an MBA focused on Branding in B2B Business (University of Wales).
Fidelis has worked with DSM for 15 years in various positions and is an experienced executive in Solutions Marketing and Sales with a demonstrated history of working in the Animal Nutrition and Health industry.

Get in touch with a dsm-firmenich Animal Nutrition & Health specialist or find contacts around the world to suit your needs.
At dsm-firmenich, we love to connect with you.
Follow us on any of the channels below.
We detected that you are visitng this page from United States. Therefore we are redirecting you to the localized version.



