Transformar una década de conocimiento y experiencia en beneficios para una producción animal sostenible

El uso de la proteasa exógena en la producción animal continúa siendo motivo de gran interés desde su introducción hace una década. La proteasa, una enzima que mejora la digestibilidad de las proteínas en el tracto digestivo de los animales, desempeña un rol fundamental en la nutrición animal. Al hacer que las proteínas sean más digeribles y estén disponibles para su absorción, las proteasas mejoran la utilización de las proteínas presentes en los alimentos balanceados, reducen su costo, disminuyen las emisiones de nitrógeno y, en general, mejoran la eficiencia y la sostenibilidad de los huevos, la carne y la producción acuícola. A lo largo de los años, la optimización de su aplicación ha mejorado a medida que avanza nuestro conocimiento sobre los efectos específicos en los diferentes ingredientes de los alimentos balanceados (tanto en digestibilidad ileal estandarizada de los aminoácidos como en respuesta energética) y los efectos adyacentes de las proteasas en la resiliencia intestinal, la función de barrera de la mucina y el transporte de nutrientes. También se ha centrado mucha atención sobre la forma en que las proteasas exógenas complementan la actividad de otras enzimas alimentarias, como las fitasas y las carbohidrasas, y la manera de combinarlas como parte de una estrategia enzimática más amplia, con valores adecuados de matriz en formulaciones de menor costo. Asimismo, se ha avanzado considerablemente en la comprensión de la enorme diversidad de la superfamilia de las proteasas, lo cual indica claramente que las características específicas de la molécula de proteasa (perfil de pH, especificidad del sustrato, inhibición potencial y estabilidad) son claves para la eficacia en el animal.
Seleccionar la proteasa ‘adecuada’ para una aplicación específica representa un enorme reto, ya que la gran mayoría de las proteasas disponibles probablemente no sean apropiadas para aplicación en la alimentación animal. La Alianza ‘dsm-firmenich | Novozymes Feed Enzyme’ cuenta con sólida experiencia en la oferta de soluciones eficaces, de alta calidad y científicamente comprobadas para el sector de la producción animal sostenible. Dicha Alianza también fue pionera en el área de proteasas exógenas para la alimentación animal con la introducción de la primera generación de proteasas (RONOZYME® ProAct) hace una década. Asimismo, la Alianza llevó a cabo un amplio y exclusivo proyecto de cribado in vivo, que culminó con el desarrollo de la segunda generación de proteasas (ProAct 360™). El objetivo del presente artículo es destacar el estatus actual y la optimización de las proteasas exógenas en el sector de enzimas nutricionales, subrayando su importancia para la mejora en la utilización de los nutrientes y la sostenibilidad en la producción de animales monogástricos. El documento también aborda los efectos directos e indirectos de las proteasas exógenas, específicamente sus interacciones con otras enzimas. En la conclusión, se destaca la necesidad de una formulación estratégica, que tenga en cuenta las diferencias de materia prima y la divergencia de aminoácidos, para maximizar el impacto positivo de las proteasas en la nutrición animal y la eficiencia de la producción.
Actualmente, el mercado de enzimas exógenas para alimentación animal representa alrededor de 1.100 millones de dólares anuales y supone un ahorro para el sector de aproximadamente 5 mil millones de dólares anuales, por menor costo de nutrientes y reducción de emisiones al medio ambiente. Además de mejorar la utilización de los nutrientes, el uso de enzimas nutricionales ejerce un impacto positivo sustancial en la sostenibilidad medioambiental, social y económica del sector de producción animal. Inicialmente, la comercialización de enzimas nutricionales se centraba en las carbohidrasas (beta-glucanasas y arabinoxilanasas), para abordar los efectos antinutricionales de los pentosanos solubles de alto peso molecular presentes en el trigo, la cebada, el centeno y el triticale, y estaba dirigido a los mercados del norte de Europa, Australia y Canadá. Paralelamente al crecimiento del sector de las carbohidrasas, surgieron las fitasas, a principios de los años 90, para reemplazar a los fosfatos inorgánicos finitos en las dietas de animales no rumiantes y reducir los efectos antinutricionales del ácido fítico. Posteriormente, el mercado de carbohidrasas se amplió con actividades adicionales como la alfa-amilasa, alfa-galactosidasa y beta-mananasa y se extendió a las dietas a base de maíz y sorgo, centrado también en las harinas de proteínas vegetales como soya y colza. Más recientemente, se ha observado una evolución en el mercado de enzimas nutricionales, con la adaptación de la nutrición de precisión con fitasa para degradar el ácido fítico de la dieta de forma más rápida y completa, y el surgimiento de un nuevo segmento de proteasas monocomponentes.
Todas estas diferentes etapas evolutivas han generado un valor excepcional para el usuario final de enzimas para alimentación animal, así como también una serie de soluciones para muchos de los retos en el área de nutrición. Sin embargo, con cada etapa de evolución también ha aumentado la complejidad, tanto en lo que se refiere al número de productos que compiten entre sí como a la evaluación adecuada del valor combinado de las enzimas nutricionales, ya sea para ahorro en los costos de alimentación o para el desempeño animal. Recientemente, se ha cuestionado la utilidad de las proteasas monocomponentes en dietas que contienen carbohidrasa y fitasa. El argumento sería que algunas dietas pueden requerir solamente uno o dos productos enzimáticos, con una sustancial canibalización del valor con cada nueva adición de enzimas.
Las proteasas, también conocidas como peptidasas o enzimas proteolíticas, representan una clase muy conocida de enzimas, aplicadas con éxito en diferentes sectores desde hace muchos años. La diversidad de proteasas es enorme y, actualmente, se encuentran disponibles públicamente más de medio millón de diferentes secuencias de proteasas (https://www.uniprot.org/).
En el contexto de las proteasas existe una enorme diversidad en términos de especificidad y estabilidad, además de perfiles de temperatura y pH. Las proteasas se clasifican en diferentes familias y subfamilias, en función de la identidad de secuencia en el sitio activo. Las seis principales familias de proteasas son: serina proteasas, metaloproteasas, cisteína proteasas, aspartato proteasas, glutámico proteasas y treonina proteasas. Al mismo tiempo, las proteasas se pueden organizar en dos grupos: exopeptidasas y endopeptidasas. Las exopeptidasas, incluidas las di y tri peptidil peptidasas, actúan sobre los extremos de las cadenas de proteínas liberando aminoácidos, dipéptidos o tripéptidos. Las endopeptidasas actúan dentro de las cadenas de proteínas liberando y solubilizando fragmentos más grandes de proteína. El sitio de clivaje depende además de la especificidad de la proteasa. En proteasas muy específicas, como la tripsina, el clivaje ocurre preferentemente próximo a aminoácidos concretos, limitando así el número de cortes en una cadena proteica, mientras que las proteasas con amplia especificidad generan una hidrólisis más extensa. La amplia especificidad de algunas proteasas exógenas representa una ventaja para su uso en la alimentación animal, ya que permite la hidrólisis proteica de una amplia gama de materias primas diferentes.
Incluso cuando se encuentra una proteasa adecuada que aparentemente cumple los criterios anteriormente mencionados, es esencial que dicha proteasa se añada al producto final en dosis suficientes para que ejerza un efecto in vivo. De acuerdo con las condiciones elegidas para medir la actividad (pH, temperatura, sustrato), la clasificación de los productos con base en la actividad de la proteasa puede variar.
Por ese motivo y para que la comparación sea útil, se prefiere que las condiciones sean fisiológicamente relevantes. Para la aplicación de proteasas en la alimentación animal, también es importante observar el perfil de actividad de la proteasa en función del pH y la temperatura, así como la estabilidad de dicha proteasa. Asimismo, es necesario optimizar la compatibilidad de la proteasa suplementada con las proteínas endógenas y de la dieta. En la Figura 1, se indican una serie de perfiles de pH para proteasas, subrayando así la gran diferencia entre las distintas proteasas.
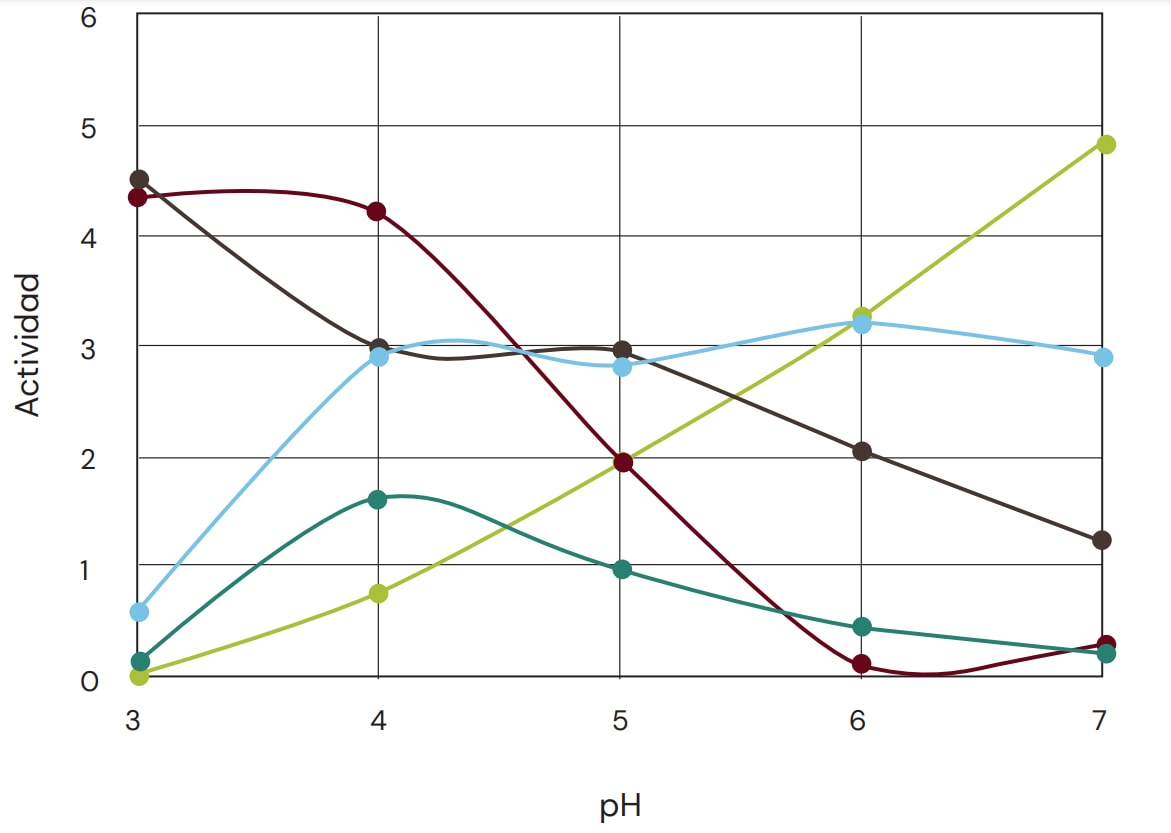 Fig. 1: Ejemplos de perfiles de actividad de pH para diferentes proteasas, destacando la diferencia significativa en perfiles de pH. Incluso en una misma familia de proteasas, los perfiles de pH pueden variar significativamente. La actividad en diferentes valores de pH se midió en una mezcla 30:70 de harina de soya/maíz, monitoreando el aumento de aminoterminales en la solución tras hidrólisis de 3 horas. Sólo se consideraron valores de pH fisiológicamente relevantes.
Fig. 1: Ejemplos de perfiles de actividad de pH para diferentes proteasas, destacando la diferencia significativa en perfiles de pH. Incluso en una misma familia de proteasas, los perfiles de pH pueden variar significativamente. La actividad en diferentes valores de pH se midió en una mezcla 30:70 de harina de soya/maíz, monitoreando el aumento de aminoterminales en la solución tras hidrólisis de 3 horas. Sólo se consideraron valores de pH fisiológicamente relevantes.
La estabilidad al peletizado es un parámetro de fundamental importancia. La mayoría de los alimentos balanceados para aves se peletizan, y las principales enzimas se suministran en forma granulada o como productos secos para mezclar con el alimento y posteriormente pasar por el proceso de peletizado. Por lo tanto, es importante que la proteasa pueda conservar su actividad tras una posible etapa de peletizado en condiciones de alta temperatura, presión y humedad. Esto suele representar un reto, dado que las enzimas en entornos biológicos no están adaptadas a estas condiciones. La estabilidad durante el peletizado se puede mejorar en algunos casos con una formulación o recubrimiento específico de los gránulos. Esto también protege a los trabajadores contra la formación de polvo, permitiendo así el uso seguro de los productos enzimáticos. En la Figura 2 se indica la estabilidad al vapor (indicador de estabilidad al peletizado) de una serie de productos comerciales que declaran tener actividad de proteasa. La mayoría de los productos, o no presentan actividad de proteasa tan alta a la dosis recomendada, o contienen proteasas que no son estables al vapor. Se puede concluir así que la calidad de los productos comerciales para alimentación animal disponibles en el mercado varía significativamente, lo cual aumenta aún más la complejidad a la hora de estimar el valor en general de las proteasas nutricionales.
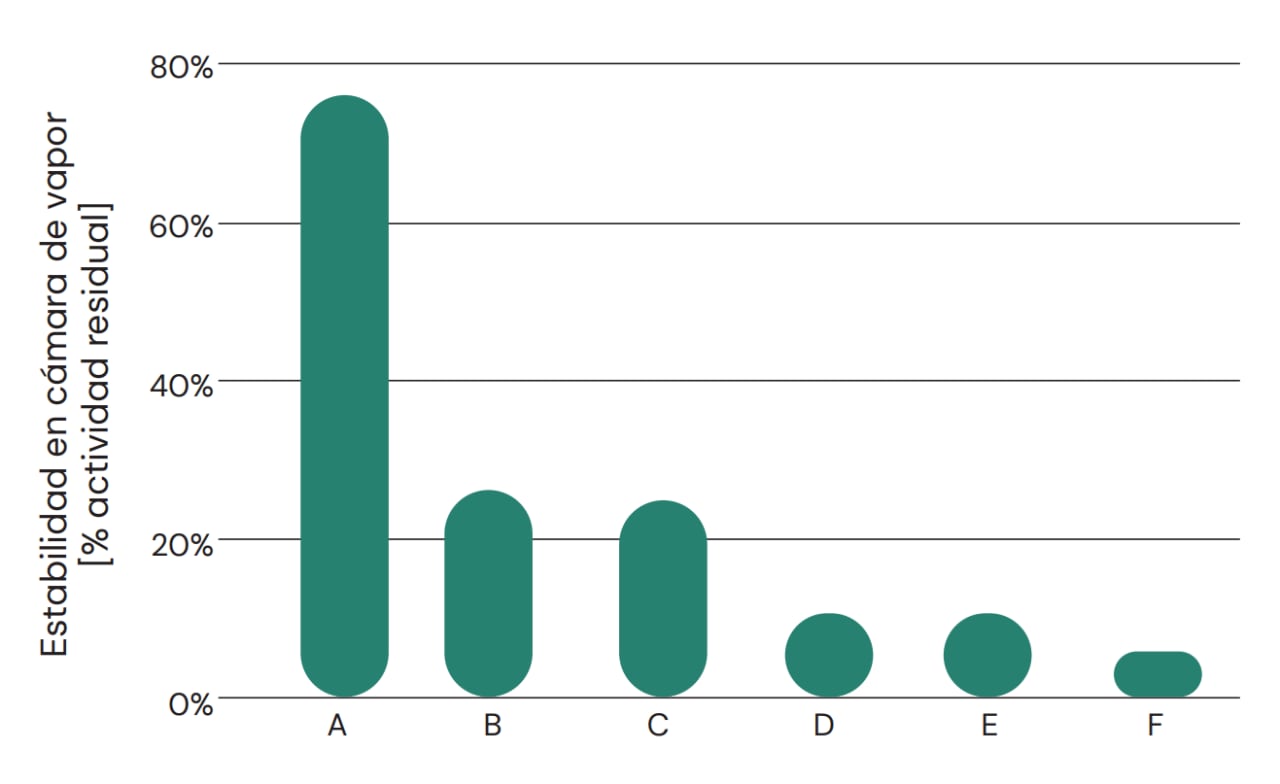 Fig. 2: Estabilidad en cámara de vapor de diferentes productos comerciales que declaran actividad de proteasa. Solamente algunos pocos productos comerciales pueden retener la actividad de proteasa tras exposición a condiciones que simulan el proceso de peletizado, en términos de alta temperatura y humedad. Los productos granulados se cocieron al vapor a 95 °C durante 90 segundos, y la actividad residual se calculó por comparación con la muestra no tratada. A = ProAct 360™, B = Competidor (B), C = Competidor (C), D = Competidor (D), E = Proteasa neutra, F = Competidor (F).
Fig. 2: Estabilidad en cámara de vapor de diferentes productos comerciales que declaran actividad de proteasa. Solamente algunos pocos productos comerciales pueden retener la actividad de proteasa tras exposición a condiciones que simulan el proceso de peletizado, en términos de alta temperatura y humedad. Los productos granulados se cocieron al vapor a 95 °C durante 90 segundos, y la actividad residual se calculó por comparación con la muestra no tratada. A = ProAct 360™, B = Competidor (B), C = Competidor (C), D = Competidor (D), E = Proteasa neutra, F = Competidor (F).
Desde el lanzamiento en 2008 de la primera proteasa para alimentos balanceados, RONOZYME® ProAct, Novozymes y dsm-firmenich han investigado de forma continua la eficacia de las proteasas en la alimentación animal. Encontrar una proteasa que funcione con mayor eficacia que RONOZYME® ProAct en los animales no ha sido tarea fácil y ha supuesto un gran esfuerzo de investigación y desarrollo. En los últimos años, más de 500 nuevas proteasas experimentales se han sometido a pruebas in vitro. Las enzimas pertenecían a diferentes familias y poseían características diferentes en lo referente al sitio de acción (endoproteasas versus exoproteasas) y a las condiciones óptimas de temperatura y pH (proteasas ácidas versus alcalinas).
A partir de un conjunto de 500 proteasas, se hicieron pruebas en animales con 71 moléculas, mediante un ensayo de cribado sensible y de alto rendimiento desarrollado por la Alianza dsm-firmenich | Novozymes. En la etapa inicial, se evaluó la digestibilidad del nitrógeno en yeyuno e íleo para obtener una imagen dinámica de su absorción y digestión. Es importante evaluar la acción de la proteasa en los diferentes segmentos del intestino, ya que ello beneficiará al animal, haciendo que los péptidos estén más fácilmente disponibles para su absorción y reduciendo el nivel de proteína no digerida que llega al intestino posterior. Estas etapas permitieron reducir el número de proteasas candidatas de 71 a 3, y finalmente, tras exhaustivos ensayos in vivo, se seleccionó ProAct 360™. Más del 80% de las proteasas examinadas in vivo no presentaron ningún efecto o, en el caso de algunas proteasas, se observó incluso un efecto negativo sobre la digestibilidad de la proteína. Tomando como base extensos ensayos y múltiples datos, se concluyó que ProAct 360™ era superior a las demás candidatas y fue así seleccionada para desarrollo posterior.
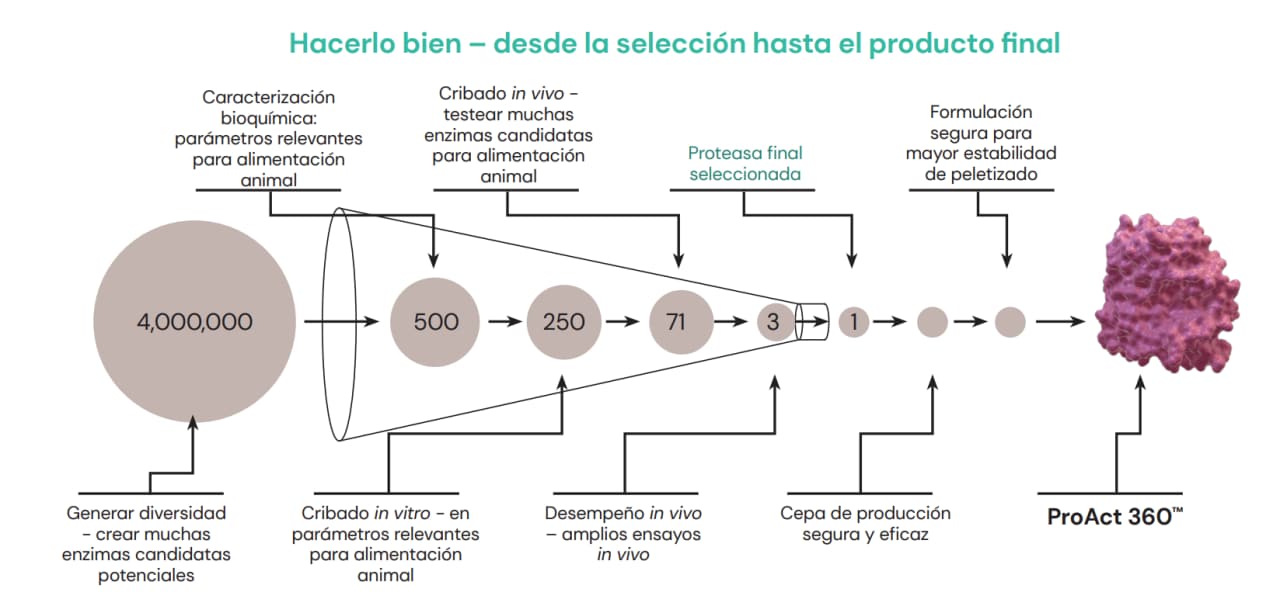 Fig. 3: Proceso de cribado amplio y exclusivo para seleccionar la mejor candidata como proteasa nutricional de segunda generación: la molécula de ProAct 360™. La Alianza utilizó toda la tecnología y el conocimiento desarrollados a lo largo de los años para generar una proteasa monocomponente, única y superior en comparación con otros productos del mercado que afirman poseer actividad de proteasa.
Fig. 3: Proceso de cribado amplio y exclusivo para seleccionar la mejor candidata como proteasa nutricional de segunda generación: la molécula de ProAct 360™. La Alianza utilizó toda la tecnología y el conocimiento desarrollados a lo largo de los años para generar una proteasa monocomponente, única y superior en comparación con otros productos del mercado que afirman poseer actividad de proteasa.
ProAct 360™ es una proteasa de la familia de las serina proteasas, subfamilia A de las subtilisinas (MEROPS ID S08.113*) con actividad de endopeptidasa y amplia especificidad. ProAct 360™ se aisló a partir de una nueva especie de Bacillus (que aún no ha sido nominada), y se considera una proteasa monocomponente con 600.000 unidades de proteasa por gramo de producto. La actividad se determina en relación con un estándar enzimático, y el resultado se expresa en NFP/g. Una unidad NFP se define como la cantidad de enzima que libera aproximadamente 1 µmol de p-nitroanilina a partir de 1 mM de sustrato (N-Succinil-Ala- Ala-Pro-Phe p-nitroanilida) por minuto en pH 9.0 y 37 °C.
La evaluación in vitro de ProAct 360™ indica claramente que se trata de una proteasa para alimentación animal altamente eficaz, utilizada para hidrolizar y solubilizar las proteínas de los alimentos balanceados.
Por ser una proteasa altamente estable a la temperatura, ProAct 360™ resiste a las rigurosas condiciones aplicadas durante el peletizado de los alimentos balanceados. ProAct 360™ ejerce su función principal en el intestino delgado del animal, y ha sido seleccionada debido a su alta actividad bajo condiciones de pH y temperatura relevantes desde el punto de vista fisiológico. ProAct 360™ es una endoproteasa con amplia especificidad y clivaje en cualquier punto de la cadena proteica, lo cual le permite actuar en una amplia gama de sustratos relevantes para la alimentación animal. Al solubilizar y dividir la proteína en fragmentos menores, ProAct 360™ actúa en sinergia con las proteasas endógenas que, posteriormente, digieren aún más la proteína del alimento permitiendo así mayor absorción por el animal.
Las pruebas in vitro indican la capacidad de ProAct 360™ de aumentar la hidrólisis proteica de varios importantes ingredientes presentes en los alimentos balanceados, por encima de las proteasas digestivas endógenas. Ver Figura 4.
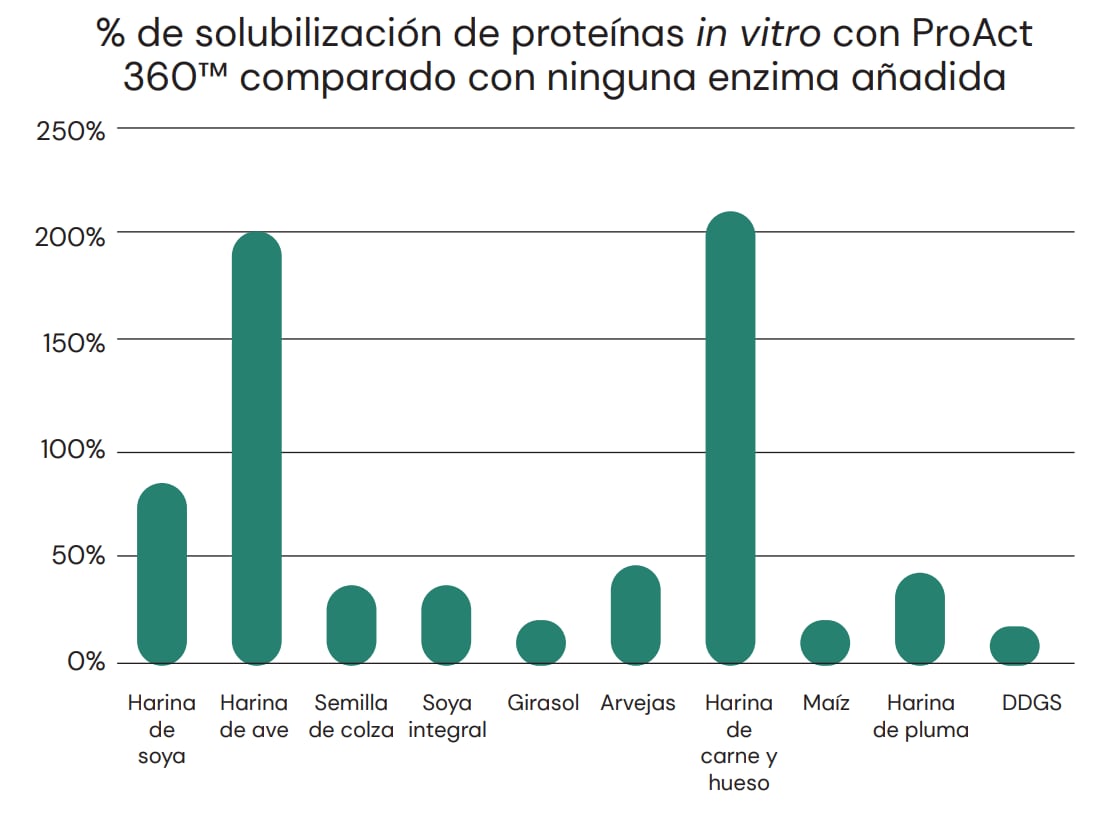 Fig. 4: Solubilización in vitro de diferentes proteínas provenientes de materias primas relevantes para la alimentación animal. Los sustratos y ProAct 360™ se incubaron durante 3 horas a 40°C y pH 5. Tras la incubación se analizó el sobrenadante mediante ensayo OPA. La solubilización de proteínas con ProAct 360™ funcionó bien en todas las materias primas de la prueba, aumentando la solubilización de proteínas entre un 16 y un 84 % (A) en sustratos vegetales y entre un 41 y un 211 % (B) en relación con la muestra control (sin enzima).
Fig. 4: Solubilización in vitro de diferentes proteínas provenientes de materias primas relevantes para la alimentación animal. Los sustratos y ProAct 360™ se incubaron durante 3 horas a 40°C y pH 5. Tras la incubación se analizó el sobrenadante mediante ensayo OPA. La solubilización de proteínas con ProAct 360™ funcionó bien en todas las materias primas de la prueba, aumentando la solubilización de proteínas entre un 16 y un 84 % (A) en sustratos vegetales y entre un 41 y un 211 % (B) en relación con la muestra control (sin enzima).
También es importante su capacidad de degradar los factores antinutricionales (FAN), por ejemplo, de la harina de soya. La harina de soya contiene diferentes factores antinutricionales como los inhibidores de tripsina y lectina, cuyo nivel depende sobre todo de las condiciones de procesamiento. Numerosos estudios han indicado que los niveles de FAN difieren en los diferentes lotes de harina de soya y que niveles crecientes de FAN acarrean consecuencias negativas en el desempeño de los animales (McNaughton et al. 1981; Palacios et al. 2004; Pacheco et al., 2014). ProAct 360™ tiene el potencial de degradar los inhibidores de tripsina y lectina de la harina de soya, como se indica en la Figura 5, colaborando así en la reducción de los efectos negativos de dichos FANs en el desempeño animal. También se ha demostrado que los inhibidores de tripsina no inhiben la acción de ProAct 360™ como ocurre con la tripsina.
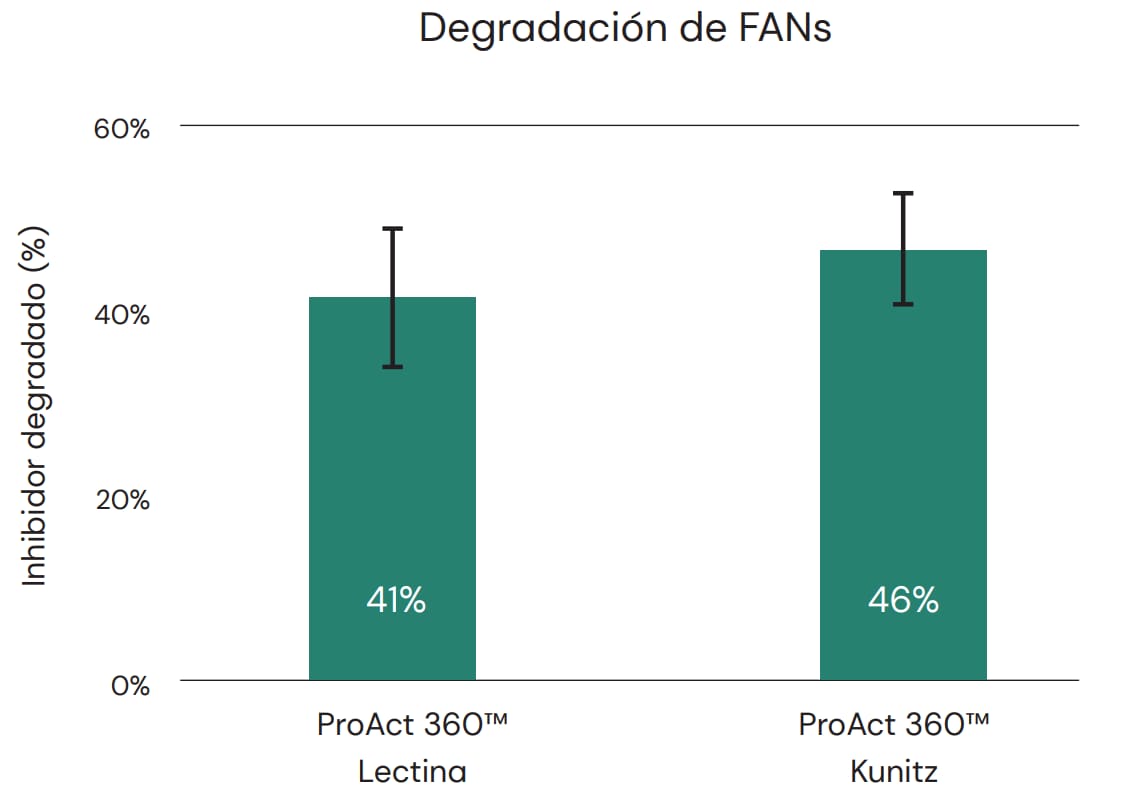 Fig. 5: ProAct 360™ tiene la capacidad de degradar inhibidores de tripsina de tipo Lectina y Kunitz de la harina de soya en geles SDS-Page. Los porcentajes indican la eficiencia de degradación calculada a partir de la intensidad de bandas. La enzima se mezcló con inhibidores de tripsina y se incubó a pH 7 y 40°C durante 2 horas.
Fig. 5: ProAct 360™ tiene la capacidad de degradar inhibidores de tripsina de tipo Lectina y Kunitz de la harina de soya en geles SDS-Page. Los porcentajes indican la eficiencia de degradación calculada a partir de la intensidad de bandas. La enzima se mezcló con inhibidores de tripsina y se incubó a pH 7 y 40°C durante 2 horas.
La evaluación in vivo indicó que el perfil de digestibilidad de los aminoácidos de ProAct 360™ es similar al de la primera generación de Ronozyme® ProAct. Sin embargo, se demostró claramente que ProAct 360™ presentaba digestibilidad de nitrógeno y aminoácidos consistentemente más alta en comparación con RONOZYME® ProAct (Fig.6). Esta mejora podría explicarse por la mayor capacidad de ProAct 360™ para aumentar la hidrólisis de proteína, contrarrestar el efecto de los factores antinutricionales y complementar las proteasas endógenas. La respuesta media para la mayor digestibilidad del nitrógeno en el caso de ProAct 360™ fue de +3.2%. Los aminoácidos relevantes en la nutrición (Met, Cys, Lys, Thr, Trp, Arg, Val, Ile y Leu) oscilaron entre +1.9% para Arg y +5.8% para Cys. La mejora en la digestibilidad del nitrógeno y de los aminoácidos puede redundar en mayor ganancia de peso y tasa de conversión alimentaria, además de mayor bienestar animal y sostenibilidad para las empresas avícolas.
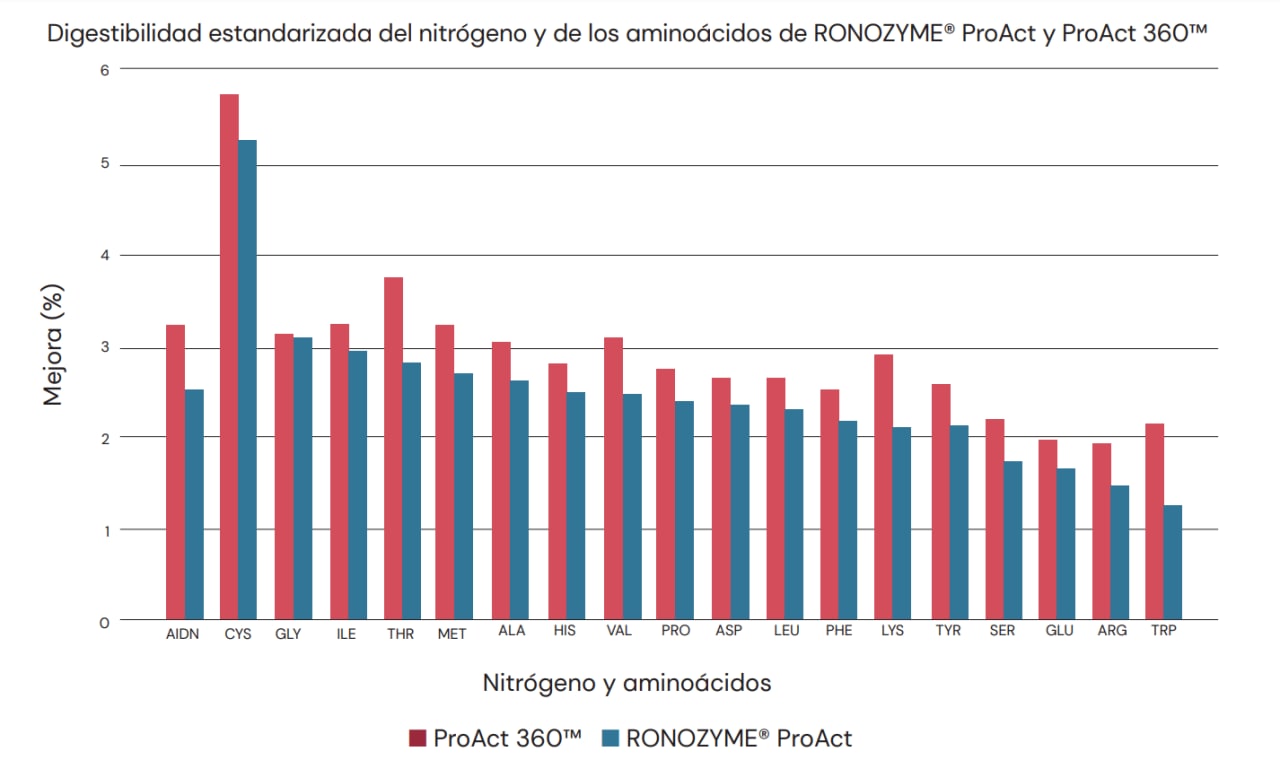 Fig. 6. Perfil de digestibilidad del nitrógeno y de los aminoácidos de RONOZYME® ProAct (azul) y ProAct 360™ (rojo).
Fig. 6. Perfil de digestibilidad del nitrógeno y de los aminoácidos de RONOZYME® ProAct (azul) y ProAct 360™ (rojo).
Asimismo, una proteasa de alta calidad se debe formular de forma que garantice un gran número de pequeños gránulos por gramo, con homogeneidad de tamaño para garantizar su miscibilidad en el alimento balanceado. Una formulación sin formación de polvo también es importante para la seguridad de los trabajadores. La formulación de ProAct 360™ se desarrolló de forma que se garanticen todos estos parámetros de seguridad y calidad, con el fin de aportar resultados consistentes y robustos en el campo (Fig. 7).
 Fig. 7: Aspecto de ProAct 360™. Los gránulos deben ser lo más pequeños y homogéneos posible para garantizar buena miscibilidad en el alimento balanceado, con bajo coeficiente de variación.
Fig. 7: Aspecto de ProAct 360™. Los gránulos deben ser lo más pequeños y homogéneos posible para garantizar buena miscibilidad en el alimento balanceado, con bajo coeficiente de variación.
Efectos directos
Como su nombre lo indica, el efecto directo de la proteasa exógena es la hidrólisis de polipéptidos de alto peso molecular en oligopéptidos de menor peso molecular (Glitsø et al., 2012). Dicho efecto depende de las características de la proteasa en cuestión, es decir, de la especificidad del sustrato, del perfil de pH y de si es una proteasa de acción endógena o exógena, como mencionado anteriormente. El sustrato para las proteasas exógenas son las proteínas no disponibles de la dieta, los antinutrientes proteicos, como los inhibidores de tripsina y lectinas, además de las proteínas en sí (donde la tasa de digestión puede aumentar en relación con la dependencia de proteasas endógenas por sí solas). La hidrólisis de estas proteínas aumenta significativamente la digestibilidad de las proteínas de la dieta (Cowieson & Roos, 2014), y la magnitud de dicho aumento depende del aminoácido, la materia prima en cuestión y la calidad inherente relativa de la proteína. Es importante resaltar que hay grandes diferencias en el efecto de la proteasa por aminoácido y también por materia prima, por lo cual es necesario prestar atención a la formulación de las dietas para acomodar los efectos de la proteasa. Es decir, valores fijos de un aminoácido o de una matriz proteica pueden generar respuestas variables en términos de desempeño.
Es importante destacar que no todas las proteasas exógenas presentan una funcionalidad favorable en las dietas de los animales no rumiantes. Cowieson y Roos (2016) han resumido los resultados de una serie de ensayos con proteasas exógenas, de origen diverso y mencionadas en la literatura científica, y se observa que muchas de ellas ejercen escasos efectos benéficos en la especie de destino. Un ejemplo concreto de lo anterior fue el trabajo desarrollado por Simbaya et al. (1996), quienes examinaron la eficacia de una serie de cinco proteasas exógenas diferentes para pollos de engorde. Dichos autores observaron que solamente una de estas proteasas tenía un efecto benéfico en el desempeño de los pollitos e, incluso en ese caso, el alcance del efecto dependía del contenido y la fuente de proteínas de la dieta a la cual se añadía. Recientemente, Walk et al. (2018) publicaron resultados similares de fracaso a la hora de demostrar la eficacia de las nuevas proteasas experimentales. Ello estaría asociado con el acceso a una diversidad limitada de candidatas adecuadas de proteasa, a una formulación de la dieta demasiado simplista (sin tener en cuenta los efectos diferenciales entre los aminoácidos y las materias primas) y al no reconocimiento de posibles diferencias en el efecto según la especie animal y la edad.
Efectos indirectos
Si bien el efecto directo de la proteasa exógena es la hidrólisis del sustrato – antinutrientes proteicos y proteína de la dieta que de otra forma no se digerirían en el intestino – hay una serie de efectos ‘extraproteicos’ significativos (no muy diferentes de los efectos extrafosfóricos de la fitasa). Dichos efectos se asocian a los beneficios de la proteasa sobre la salud intestinal (Peek et al., 2009; Kalmendahl y Tauson, 2012; Cowieson et al., 2015), y la mejora en la digestibilidad de nutrientes alternativos no proteicos, como la energía (mediada a través de la mayor digestibilidad de la grasa y el almidón).
Una preocupación habitual cuando se trata del uso de múltiples enzimas en la misma dieta es la canibalización del valor. No cabe duda de que, si bien las diferentes clases de enzimas (fitasas, carbohidrasas y proteasas) no comparten los mismos sustratos, la hidrólisis de estos diferentes sustratos genera una serie de efectos similares sobre la digestibilidad, que tienen un efecto acumulativo (aunque probablemente no totalmente aditivo) sobre el desempeño animal. Por ejemplo, se ha demostrado que las carbohidrasas (principalmente las xilanasas y glucanasas) (Cowieson, 2010); la fitasa (Cowieson et al., 2018b,c); y la proteasa (Cowieson y Roos, 2014) aumentan la digestibilidad ileal de los aminoácidos en animales no rumiantes. En el caso de la mejora de la digestibilidad de los aminoácidos con enzimas exógenas, ello se debe tanto a la reducción del flujo de aminoácidos endógenos (como ocurre principalmente en el caso de la fitasa) como también a una mejora verdadera en la digestibilidad de la proteína del alimento (como ocurre con las proteasas y las carbohidrasas). La falta de aditividad entre estas clases de enzimas está vinculada principalmente a la superposición de efecto sobre la pérdida endógena (como bien se observa en la evaluación de la digestibilidad de aminoácidos en ingredientes únicos, originando así el desarrollo del ensayo de DIE). En el caso de la fitasa, los efectos sobre la digestibilidad de los aminoácidos pueden vincularse directamente a la reducción en la pérdida de proteínas endógenas (en gran parte de mucina, pero también de pepsina). Estos efectos están relacionados con el efecto del fitato sobre la solubilidad de las proteínas en el intestino y el aumento que ello ejerce en la producción endógena de mucina y pepsina (resumen de Cowieson et al., 2009).
De hecho, Cowieson et al. (2008) observaron una correlación directa entre el efecto del fitato y la fitasa en el flujo de aminoácidos endógenos en pollos de engorde y el perfil de aminoácidos de la mucina y la pepsina. Además, la estimulación de la pérdida excesiva de aminoácidos endógenos por ingestión de ácido fítico se puede resolver fácilmente con inclusión de concentraciones relativamente bajas de fitasa microbiana, sin otros beneficios observados cuando la inclusión aumenta por encima de aproximadamente 500-750 FYT/kg (Cowieson et al., 2018b,c). Estos resultados ya se habían observado anteriormente (Shirley & Edwards, 2003; Fig. 8). Este hecho sugiere que cualquier canibalización del valor de las carbohidrasas o proteasas por la fitasa sobre la digestibilidad de los aminoácidos quedará evidente con bajas concentraciones de inclusión de fitasa, y esto no se altera a medida que se eleva la dosis de fitasa. En el caso de las carbohidrasas, los efectos sobre la digestibilidad de los aminoácidos parecen no estar muy vinculados al flujo de aminoácidos endógenos.
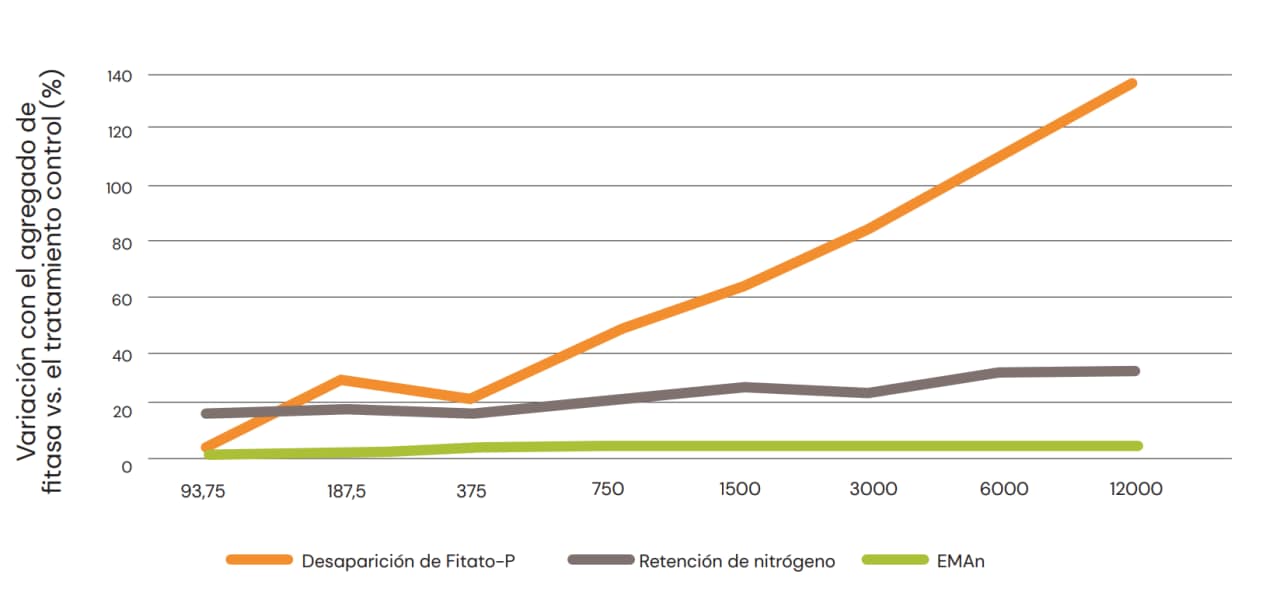 Fig. 8: Efecto del aumento de la dosis de fitasa sobre la desaparición del fitato-P, la retención de nitrógeno y la Energía Metabolizable Aparente corregida por nitrógeno (EMAn) en pollos de engorde alimentados con dieta a base de maíz/harina de soya (Shirley & Edwards, 2003).
Fig. 8: Efecto del aumento de la dosis de fitasa sobre la desaparición del fitato-P, la retención de nitrógeno y la Energía Metabolizable Aparente corregida por nitrógeno (EMAn) en pollos de engorde alimentados con dieta a base de maíz/harina de soya (Shirley & Edwards, 2003).
Esto toma como base un amplio metaanálisis, mostrando que ningún aminoácido específico se beneficia más que otro de la adición de xilanasa o glucanasa, algo que no ocurriría si el flujo de proteína endógena sufriera una influencia desproporcional en relación con la digestibilidad de la proteína alimentaria (Cowieson y Bedford, 2009). Lo anterior sugiere que el efecto de las carbohidrasas sobre la digestibilidad ileal de los aminoácidos puede en gran parte ser aditivo al efecto de la fitasa, dado que las primeras modifican la digestibilidad de la dieta (supuestamente a través de mecanismos genéricos que involucran permanencia gástrica del alimento, fermentación del intestino posterior, menor viscosidad del contenido del lumen y escape de contenido de la pared celular), al tiempo que la segunda modifica el flujo de proteínas endógenas. Por último, las proteasas proporcionan mejoras en la digestibilidad de los aminoácidos, por un lado, reduciendo la pérdida de mucina endógena (aproximadamente el 30% de su efecto), y, por otro, aumentando la retención de aminoácidos de la dieta (Cowieson & Roos, 2016). Por lo tanto, se puede prever cierta merma del efecto de la fitasa sobre la digestibilidad de los aminoácidos mediante la adición de proteasa, especialmente para los aminoácidos que se encuentran comúnmente en la proteína endógena, por ejemplo, Thr, Cys, Gly, Ser y Pro.
La soya almacena proteínas en vacuolas de almacenamiento de proteínas (PSV según su sigla en inglés), que son estructuras globulares compuestas principalmente de proteínas, ácido fítico y minerales (Jiang et al., 2001). El ácido fítico se concentra en partes densas de las PSVs denominadas globoides de fitato. Dado que el fitato presente en estos globoides puede estar incorporado a las proteínas, no se espera que esté fácilmente accesible para desfosforilación por una fitasa. Bohn et al. (2007) demostraron que la fitasa degrada el fitato libre y soluble mucho más rápido que el fitato presente en los globoides de fitato. Las proteínas del globoide pueden retardar la acción de la fitasa, bloqueando el acceso al ácido fítico. Se ha demostrado que ProAct 360™ degrada las proteínas presentes en las vacuolas de almacenamiento de proteínas (PSVs), lo cual lleva a una mayor solubilización del ácido fítico que, de ese modo, puede ser fácilmente desfosforilado por la fitasa.
El efecto de ProAct 360™ sobre las vacuolas de almacenamiento (PSVs) de la soya se indica en la Figura 9. Se fijaron secciones de tejido de cotiledón de soya, se deshidrataron y se incubaron con ProAct 360™, y el efecto de la proteasa se visualizó mediante microscopía electrónica de barrido. Se observaron numerosos orificios en las vacuolas de almacenamiento de proteínas del tejido tratado con la enzima, lo cual demuestra la degradación de la proteína presente en las PSVs.
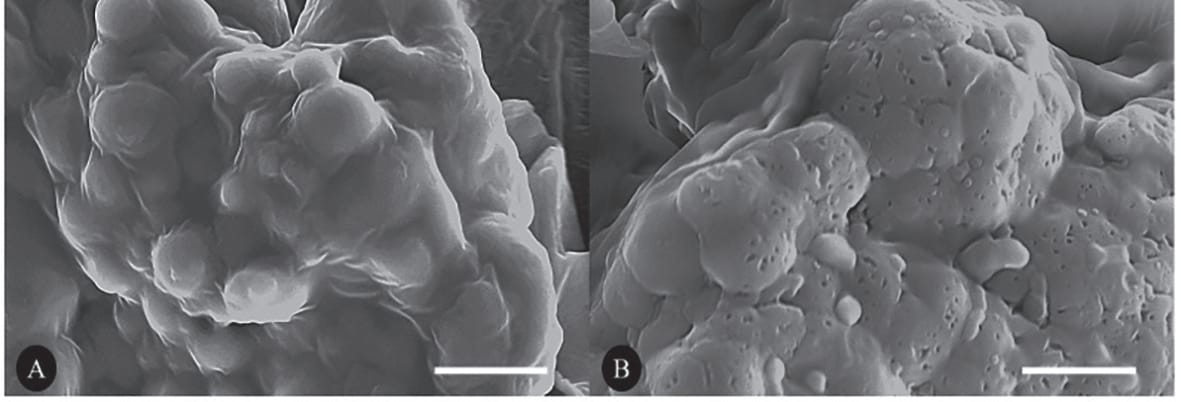 Fig. 9: Secciones de tejido fijado de cotiledón de soya no tratado/tratado con ProAct360™ y visualizado mediante microscopía electrónica de barrido. Se observan numerosos orificios en las vacuolas de almacenamiento de proteína del tejido tratado con enzima. (A) Control sin proteasa; (B) con adición de proteasa.
Fig. 9: Secciones de tejido fijado de cotiledón de soya no tratado/tratado con ProAct360™ y visualizado mediante microscopía electrónica de barrido. Se observan numerosos orificios en las vacuolas de almacenamiento de proteína del tejido tratado con enzima. (A) Control sin proteasa; (B) con adición de proteasa.
Considerar las diferencias de materias primas
No hay duda de que estas enzimas no influyen en todas las fuentes de proteínas en la misma medida (Figura 10). Esto probablemente está vinculado con la especificidad de una cierta proteasa para diferentes tipos de proteínas, como proteínas hidrófobas o polares, cargas variables, secuencias de aminoácidos, entre otros factores, así como la digestibilidad inherente de los aminoácidos provenientes de diferentes materias primas.
Esto también ocurre en el caso del conjunto de proteasas endógenas, donde se ha demostrado que algunas proteínas alimentarias escapan en gran parte a la digestión. Por ejemplo, Cowieson et al. (2017) utilizaron la proteómica para determinar el origen de los péptidos en el íleon de pollos de engorde alimentados con dieta a base de maíz/soya y observaron que la mayoría de los fragmentos peptídicos procedían de proteínas de almacenamiento no digeridas de harina de soya. Por lo tanto, no sorprende que las proteasas exógenas específicas también 'favorezcan' algunas fuentes de proteínas más que otras y, por lo tanto, presenten compatibilidad variable con las diferentes materias primas del alimento. Se puede optimizar el efecto de la proteasa in vivo haciendo una evaluación cuidadosa de la mezcla de la dieta (fuente y proporcionalidad de la proteína de la dieta) para elaborar una matriz de respuesta adecuada para esa dieta específica.
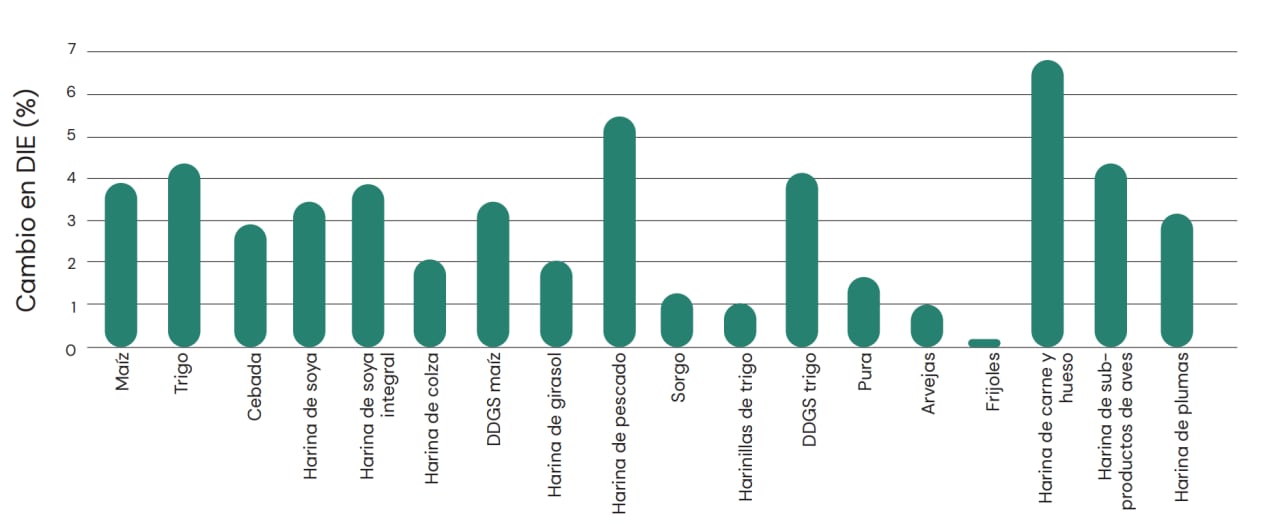 Fig. 10: Efecto de ProAct 360™ sobre la digestibilidad ileal estandarizada de aminoácidos (DIE; promedio de 9 aminoácidos) en ingredientes habituales de alimento balanceado. DDGS maíz= granos de destilería de maíz con solubles; DDGS trigo= granos de destilería de trigo con solubles.
Fig. 10: Efecto de ProAct 360™ sobre la digestibilidad ileal estandarizada de aminoácidos (DIE; promedio de 9 aminoácidos) en ingredientes habituales de alimento balanceado. DDGS maíz= granos de destilería de maíz con solubles; DDGS trigo= granos de destilería de trigo con solubles.
Considerar la divergencia entre aminoácidos
La formulación de las dietas para animales no rumiantes depende en gran medida de mantener la densidad adecuada de aminoácidos y también las proporciones con respecto a la lisina, para garantizar un aporte equilibrado de aminoácidos que favorezcan el crecimiento. Como las proteasas exógenas no liberan aminoácidos en la misma proporción (Figura 6), es fundamental, para que los efectos beneficiosos sobre la digestibilidad redunden en ganancias mensurables de desempeño, que la formulación con proteasas se elabore reconociendo las respuestas divergentes de los aminoácidos. Dichas respuestas son específicas para los ingredientes individuales de los alimentos balanceados, pero también lo son para aminoácidos específicos. En general, en lo que se refiere a la lisina (Lys), las mejoras asociadas con la proteasa son más sustanciales para Thr, Cys, Gly, Pro, Ala y Val y menos sustanciales para Glu, Trp, Phe, His, Ile, Met, Tyr, Leu, Arg, Asp y Ser (Fig. 11). Esto significa que el uso de proteasa puede alterar las proporciones ideales de aminoácidos en una dieta en particular. Y, de hecho, lo hará, si se aplica una matriz plana para la proteasa, que no reconozca las diferentes magnitudes de respuesta por aminoácido. Por lo tanto, es fundamental que la formulación de dietas que incluyan el uso de proteasas no se realice de forma simplista o desconsiderando la influencia de una combinación determinada de materias primas, además de la densidad y el equilibrio de los aminoácidos.
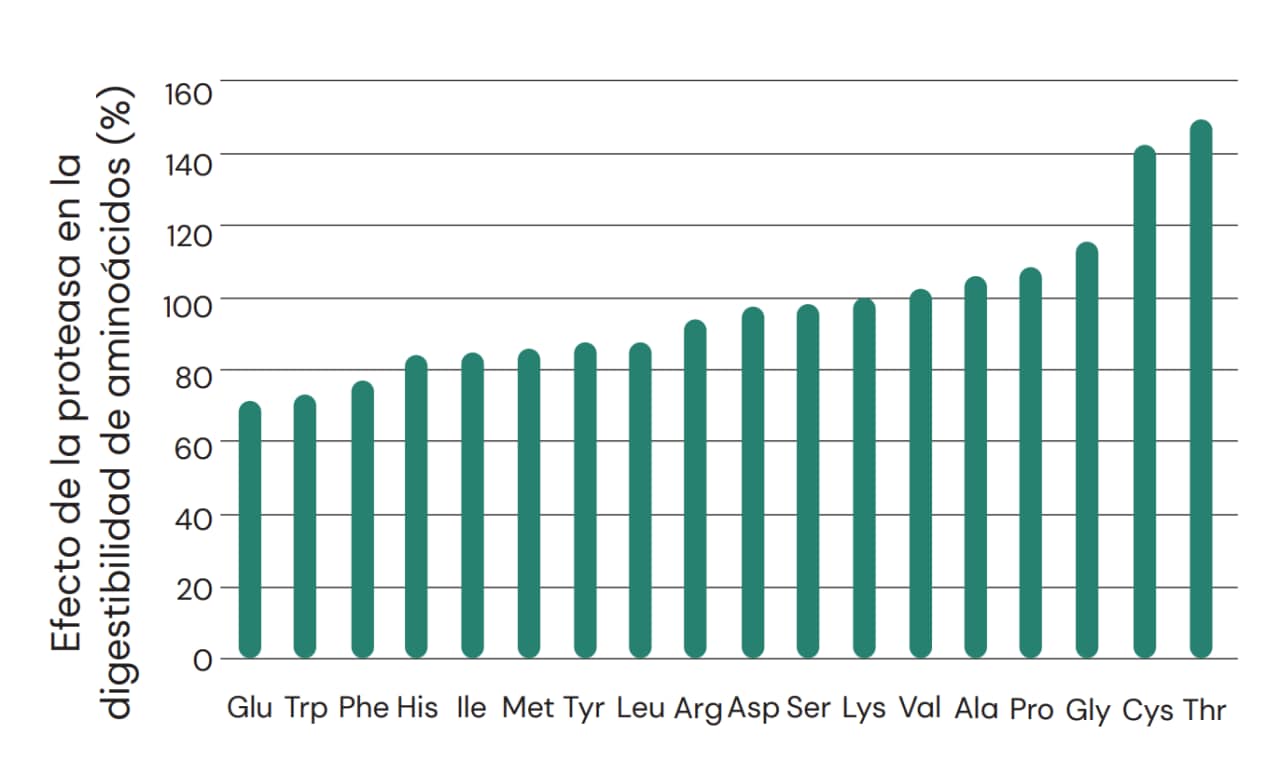 Fig. 11: Efecto de ProAct 360™ en la digestibilidad ileal de aminoácidos en animales no rumiantes expresada en relación con lisina (Lys) (Cowieson y Roos, 2014).
Fig. 11: Efecto de ProAct 360™ en la digestibilidad ileal de aminoácidos en animales no rumiantes expresada en relación con lisina (Lys) (Cowieson y Roos, 2014).
Factores adyacentes
Además del efecto de la proteasa exógena sobre materias primas y aminoácidos específicos, hay una serie de otras consideraciones que pueden influir en la magnitud y consistencia del efecto (para una revisión detallada, consultar Cowieson y Roos, 2016). Uno de estos factores es la calidad de la proteína en una materia prima determinada. Por ejemplo, un lote de harina de soya con una digestibilidad inherentemente baja presentará una respuesta elevada a la proteasa, en comparación con un lote de mayor calidad. Se ha demostrado de forma concluyente que los efectos de la proteasa son sustancialmente mayores y más consistentes cuando la digestibilidad ileal de los aminoácidos en una dieta o materia prima específica presenta valores inferiores al 90% en comparación con valores superiores (Cowieson y Roos, 2014), lo cual ofrece margen considerable para una intervención estratégica (Figura 12). Por ejemplo, los factores capaces de promover o degradar la digestibilidad de las proteínas pueden estar conceptualmente vinculados al uso de proteasa exógena, por ejemplo, especie animal, edad, procesamiento del alimento, tamaño de la partícula, etc. En ese sentido es importante señalar que la elaboración de una dieta de 'control negativo', para examinar el efecto de la proteasa exógena mediante la retirada de la proteína del alimento balanceado, muy probablemente sea algo desventajoso, dado que: a) se elimina el sustrato; b) se pierde el equilibrio de aminoácidos; c) el animal reacciona a la insuficiencia proteica regulando al alza los transportadores de péptidos, lo cual aumenta la digestibilidad de la proteína restante en la dieta. Además, es fundamental garantizar un aporte adecuado de nutrientes limitantes, como energía y fósforo, para asegurar que no se pierda el efecto benéfico de la proteasa.
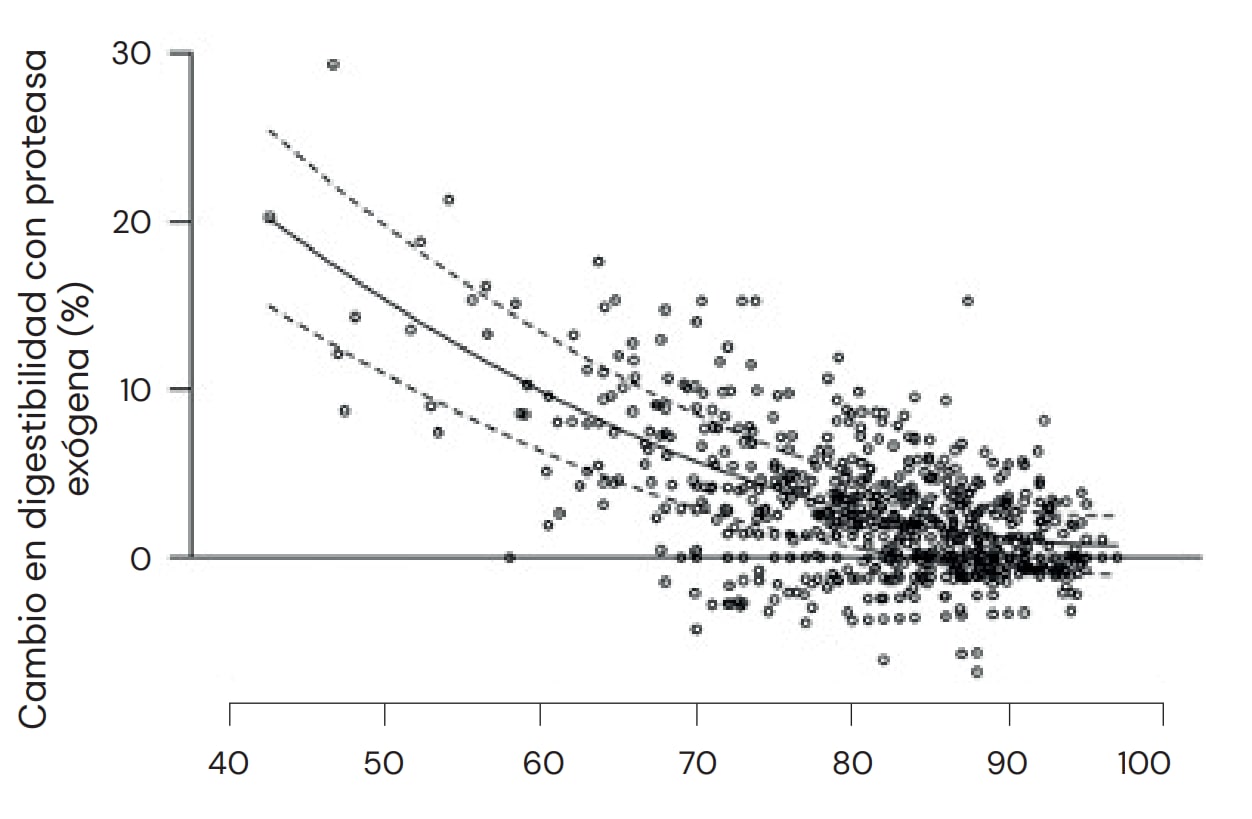 Fig. 12: Los efectos de ProAct360™ son sustancialmente mayores y más consistentes cuando la digestibilidad ileal de los aminoácidos en una dieta/materia prima específica es inferior al 90%.
Fig. 12: Los efectos de ProAct360™ son sustancialmente mayores y más consistentes cuando la digestibilidad ileal de los aminoácidos en una dieta/materia prima específica es inferior al 90%.
El desarrollo de proteasas exógenas que sean funcionales en todos los animales monogástricos, cerdos, aves y especies acuáticas, es algo sumamente difícil. Muchas de estas enzimas no funcionan, o no funcionan bien, y carecen de compatibilidad con las enzimas endógenas o con el tipo de sustratos presentes en los ingredientes habituales de los alimentos balanceados. Asimismo, una vez identificada una proteasa adecuada, no se puede simplemente adicionarla a una dieta y observar directamente respuestas útiles en lo que se refiere al desempeño. Por el contrario, es necesaria una labor considerable para mapear el efecto de la proteasa en múltiples materias primas y aminoácidos, de forma que se garantice una formulación estratégica de la dieta. También es importante destacar que el efecto benéfico de las proteasas exógenas no se limita a la mejora en la digestibilidad de los aminoácidos, sino que también se extiende a la distribución/eficiencia energética, la sostenibilidad medioambiental, la salud intestinal y otros importantes parámetros de producción.
Por último, si bien se puede prever una pequeña merma en el beneficio de los aminoácidos cuando se añaden varias enzimas a la misma dieta, es probable que dichos efectos sean moderados y orientados específicamente a los aminoácidos encontrados en la proteína endógena, por ejemplo, la treonina (Thr). Se puede concluir así que las proteasas exógenas son eficaces para mejorar la sostenibilidad de los sistemas de producción de animales no rumiantes. Sin embargo, las proteasas exógenas no se pueden utilizar de manera simplista, sin antes contar con los conocimientos básicos para utilizarlas con eficacia. Una vez sentadas dichas bases, las proteasas exógenas ofrecen un potencial significativo en la mejora de la rentabilidad de la avicultura y la porcicultura a escala mundial, además del mantenimiento de la salud y el bienestar de los animales, ante los importantes retos asociados a la alimentación de una creciente población mundial.
2 abril 2024
We detected that you are visitng this page from United States. Therefore we are redirecting you to the localized version.
