-
Latin America/PT
- Global
- América do Norte
- América Latina
Transformar uma década de conhecimento e experiência em benefícios para uma produção animal sustentável.

O uso de protease exógena na produção animal continua sendo de grande interesse desde sua introdução há uma década. A protease, uma enzima que melhora a digestibilidade das proteínas no trato digestivo dos animais, desempenha um papel fundamental na nutrição animal. Ao tornar as proteínas mais digeríveis e disponíveis para absorção, as proteases melhoram a utilização das proteínas nas rações, reduzem seu custo, reduzem as emissões de nitrogênio e, em geral, melhoram a eficiência e a sustentabilidade dos ovos, da carne e dos produtos aquícolas. Ao longo dos anos, a otimização de sua aplicação melhorou à medida que avançou nosso conhecimento sobre os efeitos específicos em diferentes ingredientes nutricionais (tanto na digestibilidade ileal padronizada de aminoácidos quanto na resposta energética) e sobre os efeitos adjacentes das proteases na resiliência intestinal, na função de barreira da mucina e no transporte de nutrientes. Muita atenção também se concentrou em como as proteases exógenas complementam a atividade de outras enzimas dietéticas, como fitases e carboidrases, e como elas podem ser combinadas como parte de uma estratégia enzimática mais ampla, com valores de matriz apropriados em formulações de baixo custo. Também houve um progresso considerável na compreensão da enorme diversidade da superfamília de proteases, indicando claramente que as características específicas da molécula de protease (perfil de pH, especificidade de substrato, inibição potencial e estabilidade) são fundamentais para a eficácia no animal.
Selecionar a protease "certa" para uma aplicação específica é um grande desafio, pois a grande maioria das proteases disponíveis provavelmente não são adequadas para aplicações em rações. A aliança entre a dsm-firmenich ǀ Novonesis para enzimas nutricionais tem uma sólida experiência no fornecimento de soluções eficazes, de alta qualidade e cientificamente comprovadas para o setor de produção animal sustentável. A Alliance também foi pioneira na área de proteases exógenas para rações, com a introdução da primeira geração de proteases (RONOZYME® ProAct) há uma década. A Alliance também realizou um extenso e exclusivo projeto de triagem in vivo, que culminou com o desenvolvimento da segunda geração de proteases (ProAct 360™). O objetivo deste artigo é destacar o status atual e a otimização de proteases exógenas no setor de enzimas para rações, enfatizando sua importância para melhorar a utilização de nutrientes e a sustentabilidade na produção de animais monogástricos. O artigo também aborda os efeitos diretos e indiretos das proteases exógenas, especificamente suas interações com outras enzimas. Na conclusão, ele destaca a necessidade de formulação estratégica, levando em conta as diferenças de matéria-prima e a divergência de aminoácidos, para maximizar o impacto positivo das proteases na nutrição animal e na eficiência da produção.
Atualmente, o mercado de enzimas exógenas para rações vale cerca de US$ 1,1 bilhão por ano e economiza para o setor aproximadamente US$ 5 bilhões por ano em termos de menores custos de nutrientes e menos emissões para o meio ambiente. Além de melhorar a utilização dos nutrientes, o uso de enzimas nutricionais tem um impacto positivo substancial na sustentabilidade ambiental, social e econômica do setor de produção animal. Inicialmente, a comercialização de enzimas nutricionais se concentrou em carboidrases (beta-glucanases e arabino-xilanases), para lidar com os efeitos antinutricionais das pentosanas solúveis de alto peso molecular no trigo, na cevada, no centeio e no triticale, e foi direcionada aos mercados do norte da Europa, da Austrália e do Canadá. Paralelamente ao crescimento do setor de carboidrases, as fitases surgiram no início dos anos 90 para substituir fosfatos inorgânicos finitos em dietas de não ruminantes e reduzir os efeitos antinutricionais do ácido fítico. Posteriormente, o mercado de carboidrases foi ampliado com atividades adicionais, como alfa-amilase, alfa-galactosidase e beta-mananase, e estendido para dietas à base de milho e sorgo, com foco também em farelos de proteína vegetal, como soja e colza. Mais recentemente, foi observada uma evolução no mercado de enzimas nutricionais, com a adaptação da nutrição de precisão com fitase para degradar o ácido fítico da dieta de forma mais rápida e completa, e o surgimento de um novo segmento de proteases monocomponentes. Todos esses diferentes estágios de evolução geraram um valor excepcional para o usuário final de enzimas nutricionais, bem como uma série de soluções para muitos dos desafios na área de nutrição. Entretanto, com cada estágio de evolução, a complexidade também aumentou, tanto em termos do número de produtos concorrentes quanto da avaliação adequada do valor combinado das enzimas nutricionais para a economia de custos de ração ou para o desempenho animal. Recentemente, a utilidade das proteases monocomponentes em dietas contendo carboidrase e fitase foi questionada. O argumento seria que algumas dietas podem exigir apenas um ou dois produtos enzimáticos, com uma canibalização substancial do valor a cada nova adição de enzima.
As proteases, também conhecidas como peptidases ou enzimas proteolíticas, representam uma classe bem conhecida de enzimas, que têm sido aplicadas com sucesso em diferentes setores há muitos anos. A diversidade de proteases é enorme e, atualmente, mais de meio milhão de sequências de proteases diferentes estão disponíveis publicamente (https://www.uniprot.org/).
No contexto das proteases, há uma enorme diversidade em termos de especificidade e estabilidade, bem como de perfis de temperatura e pH. As proteases são classificadas em diferentes famílias e subfamílias, dependendo da identidade da sequência do local ativo. As seis principais famílias de proteases são: serina proteases, metaloproteases, cisteína proteases, aspartato proteases, proteases glutâmicas e treonina proteases (Barrett et al. 2012). Ao mesmo tempo, as proteases podem ser organizadas em dois grupos: exopeptidases e endopeptidases. As exopeptidases, incluindo as di- e tri-peptidil peptidases, atuam nas extremidades das cadeias proteicas, liberando aminoácidos, dipeptídeos ou tripeptídeos. As endopeptidases atuam dentro das cadeias proteicas, liberando e solubilizando fragmentos proteicos maiores. O local de clivagem também depende da especificidade da protease. Em proteases muito específicas, como a tripsina, a clivagem ocorre preferencialmente perto de aminoácidos específicos, limitando assim o número de cortes em uma cadeia proteica, enquanto as proteases com ampla especificidade geram uma hidrólise mais extensa. A ampla especificidade de algumas proteases exógenas representa uma vantagem para seu uso na alimentação animal, pois permite a hidrólise de proteínas de uma ampla variedade de matérias-primas diferentes.
Mesmo quando se encontra uma protease adequada que aparentemente atende aos critérios acima, é essencial que essa protease seja adicionada ao produto final em doses suficientes para exercer um efeito in vivo. De acordo com as condições escolhidas para medir a atividade (pH, temperatura, substrato), a classificação dos produtos com base na atividade da protease pode variar.
Por esse motivo, condições fisiologicamente relevantes são preferíveis para uma comparação útil. Para a aplicação de proteases na alimentação animal, também é importante observar o perfil de atividade da protease em função do pH e da temperatura, bem como a estabilidade da protease. Também é necessário otimizar a compatibilidade da protease suplementada com as proteínas endógenas e dietéticas. A Figura 1 mostra uma série de perfis de pH para proteases, destacando a grande diferença entre as várias proteases.
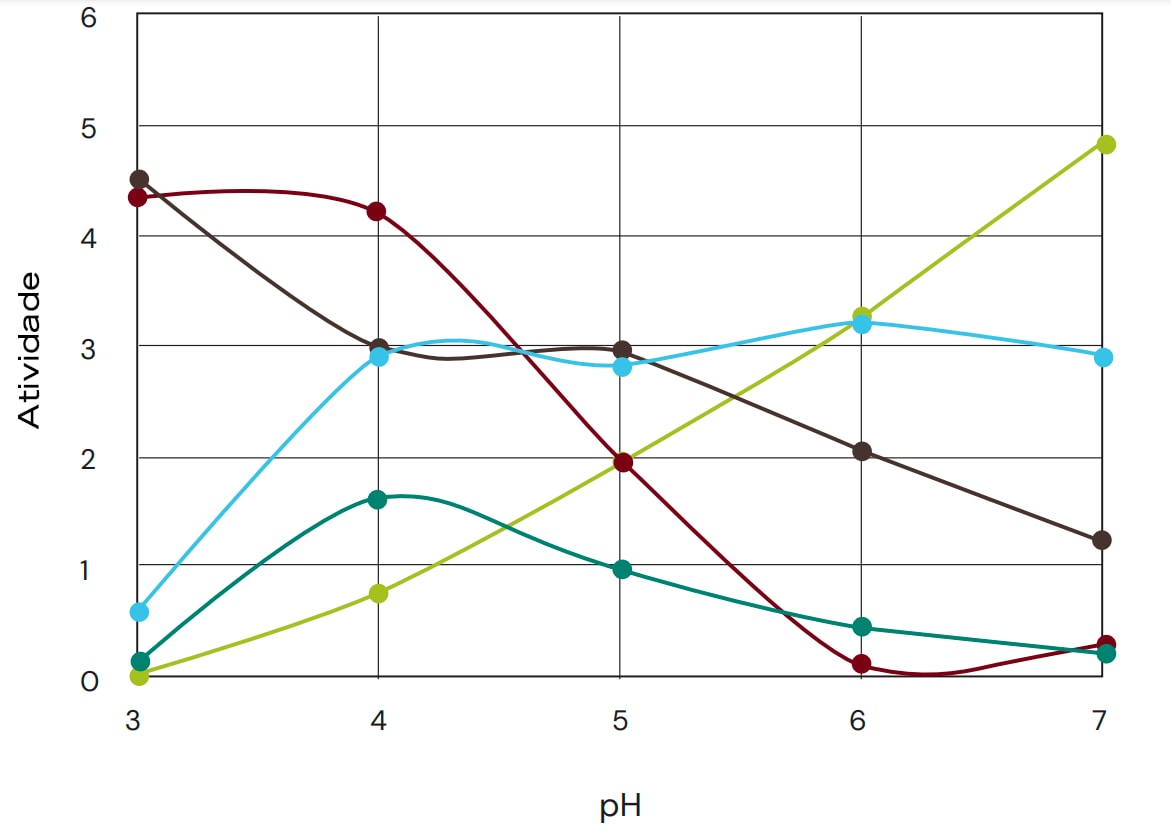 Fig. 1: Exemplos de perfis de atividade de pH para diferentes proteases, destacando a diferença significativa nos perfis de pH. Mesmo dentro da mesma família de proteases, os perfis de pH podem variar significativamente. A atividade em diferentes valores de pH foi medida em uma mistura de 30:70 de farelo de soja e milho, monitorando o aumento de amino terminais na solução após 3 horas de hidrólise. Foram considerados apenas valores de pH fisiologicamente relevantes.
Fig. 1: Exemplos de perfis de atividade de pH para diferentes proteases, destacando a diferença significativa nos perfis de pH. Mesmo dentro da mesma família de proteases, os perfis de pH podem variar significativamente. A atividade em diferentes valores de pH foi medida em uma mistura de 30:70 de farelo de soja e milho, monitorando o aumento de amino terminais na solução após 3 horas de hidrólise. Foram considerados apenas valores de pH fisiologicamente relevantes.
A estabilidade da peletização é um parâmetro extremamente importante. A maioria das rações para aves é peletizada, e as principais enzimas são fornecidas na forma granulada ou como produtos secos a serem misturados com a ração e posteriormente peletizados. Portanto, é importante que a protease possa manter sua atividade após uma possível etapa de peletização sob condições de alta temperatura, pressão e umidade. Isso geralmente é um desafio, pois as enzimas em ambientes biológicos não são adaptadas a essas condições. A estabilidade durante a peletização pode ser melhorada em alguns casos com uma formulação específica ou revestimento dos grânulos. Isso também protege os trabalhadores contra a formação de pó, permitindo assim o uso seguro dos produtos enzimáticos. A Figura 2 mostra a estabilidade ao vapor (indicador de estabilidade de peletização) de vários produtos comerciais que alegam ter atividade de protease. A maioria dos produtos não tem uma atividade de protease tão alta na dosagem recomendada ou contém proteases que não são estáveis ao vapor. Portanto, pode-se concluir que a qualidade dos produtos comerciais para ração disponíveis no mercado varia significativamente, o que aumenta ainda mais a complexidade da estimativa do valor geral das proteases nutricionais.
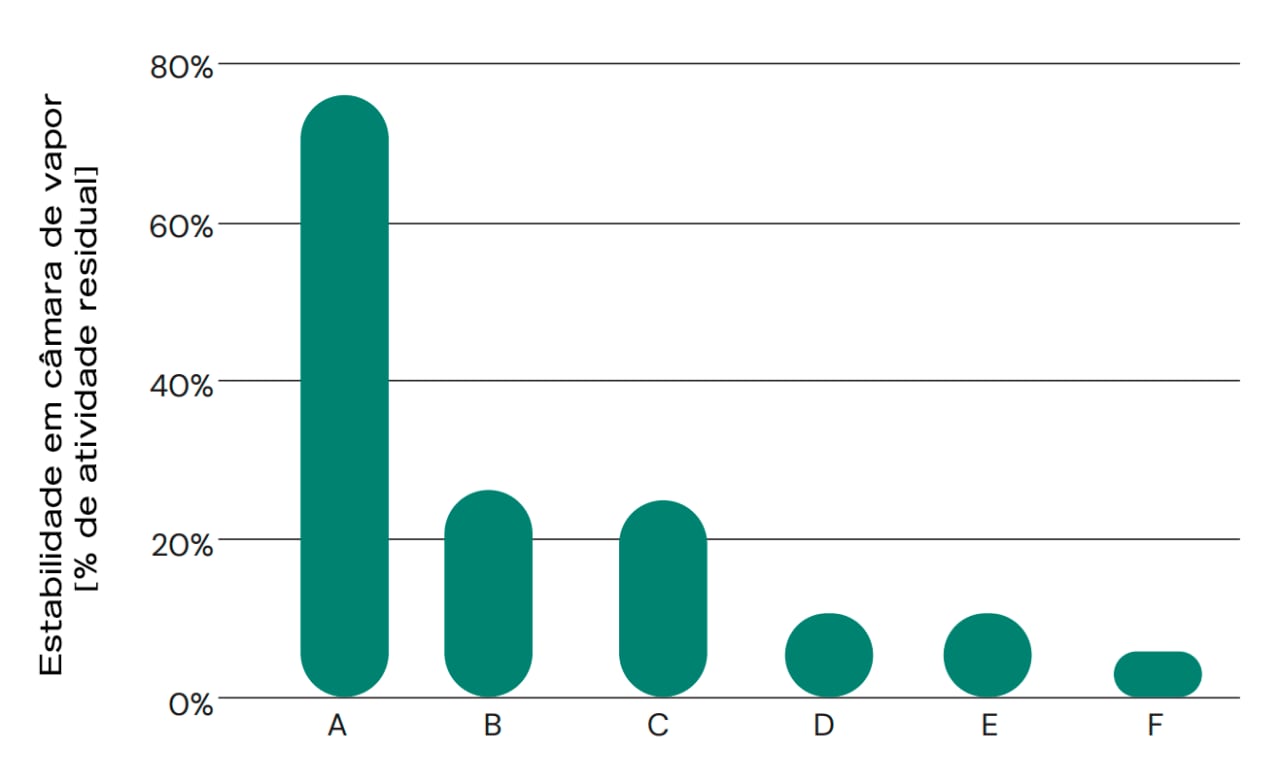 Fig. 2: Estabilidade em câmara de vapor de diferentes produtos comerciais que alegam atividade de protease. Apenas alguns produtos comerciais podem reter a atividade da protease após a exposição a condições que simulam o processo de peletização, em termos de alta temperatura e umidade. Os produtos granulados foram cozidos no vapor a 95 °C por 90 segundos, e a atividade residual foi calculada por comparação com a amostra não tratada. A = ProAct 360™, B = concorrente (B), C = concorrente (C), D = concorrente (D), E = protease neutra, F = concorrente (F).
Fig. 2: Estabilidade em câmara de vapor de diferentes produtos comerciais que alegam atividade de protease. Apenas alguns produtos comerciais podem reter a atividade da protease após a exposição a condições que simulam o processo de peletização, em termos de alta temperatura e umidade. Os produtos granulados foram cozidos no vapor a 95 °C por 90 segundos, e a atividade residual foi calculada por comparação com a amostra não tratada. A = ProAct 360™, B = concorrente (B), C = concorrente (C), D = concorrente (D), E = protease neutra, F = concorrente (F).
Desde o lançamento em 2008 da primeira protease para rações, o RONOZYME® ProAct, a Novonesis e a dsm-firmenich têm pesquisado continuamente a eficácia das proteases na alimentação animal. Encontrar uma protease que funcione de forma mais eficaz do que o RONOZYME®ProAct em animais não tem sido uma tarefa fácil e envolveu uma grande quantidade de pesquisa e desenvolvimento. Nos últimos anos, mais de 500 novas proteases experimentais foram testadas in vitro. As enzimas pertenciam a diferentes famílias e tinham características diferentes em termos de sítio de ação (endoproteases versus exoproteases) e condições ideais de temperatura e pH (proteases ácidas versus alcalinas).
De um conjunto de 500 proteases, 71 moléculas foram testadas em animais usando um ensaio de triagem sensível e de alto rendimento desenvolvido pela aliança dsm-firmenich ǀ Novonesis. No estágio inicial, a digestibilidade do nitrogênio no jejuno e no íleo foi avaliada para obter um quadro dinâmico de sua absorção e digestão. É importante avaliar a ação da protease nos diferentes segmentos do intestino, pois isso beneficiará o animal ao tornar os peptídeos mais prontamente disponíveis para absorção e reduzir o nível de proteína não digerida que chega ao intestino grosso. Essas etapas permitiram que o número de proteases candidatas fosse reduzido de 71 para 3 e, finalmente, após extensos testes in vivo, o ProAct 360™ foi selecionado (Figura 3). Mais de 80% das proteases testadas in vivo não apresentaram efeito ou, no caso de algumas proteases, foi observado até mesmo um efeito negativo na digestibilidade da proteína. Com base em testes extensivos e dados múltiplos, concluiu-se que o ProAct 360™ era superior aos outros candidatos e, portanto, foi selecionado para desenvolvimento posterior.
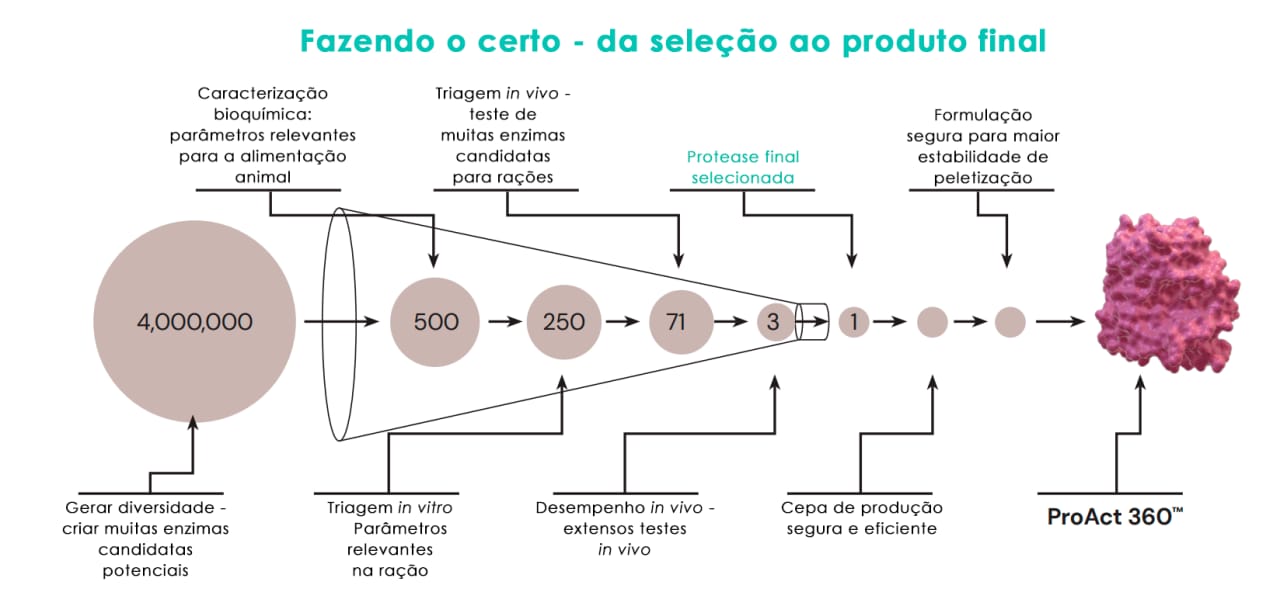 Fig. 3: Processo de triagem extenso e exclusivo para selecionar o melhor candidato como protease nutricional de segunda geração: a molécula ProAct 360™. A Alliance usou toda a tecnologia e o conhecimento desenvolvidos ao longo dos anos para gerar uma protease de componente único que é exclusiva e superior em comparação com outros produtos no mercado que alegam atividade de protease.
Fig. 3: Processo de triagem extenso e exclusivo para selecionar o melhor candidato como protease nutricional de segunda geração: a molécula ProAct 360™. A Alliance usou toda a tecnologia e o conhecimento desenvolvidos ao longo dos anos para gerar uma protease de componente único que é exclusiva e superior em comparação com outros produtos no mercado que alegam atividade de protease.
O ProAct 360™ é uma protease da família das serina proteases, subfamília A das subtilisinas (MEROPS ID S08.113*) com atividade de endopeptidase e ampla especificidade. O ProAct 360™ foi isolado de uma nova espécie de Bacillus (ainda não nomeada) e é considerado uma protease monocomponente com 600.000 unidades de protease por grama de produto. A atividade é determinada em relação a um padrão de enzima, e o resultado é expresso como NFP/g. Uma unidade NFP é definida como a quantidade de enzima que libera aproximadamente 1 µmol de p-nitroanilina de 1 mM de substrato (N-Succinyl-Ala- Ala-Pro-Phe p-nitroanilide) por minuto em pH 9,0 e 37 °C.
A avaliação in vitro do ProAct 360™ indica claramente que ele é uma protease nutricional altamente eficiente usada para hidrolisar e solubilizar proteínas em rações animais.
Por ser uma protease altamente estável à temperatura, o ProAct 360™ suporta as condições rigorosas aplicadas durante a peletização de rações. O ProAct 360™ exerce sua principal função no intestino delgado do animal e foi selecionado devido à sua alta atividade em condições de pH e temperatura fisiologicamente relevantes. O ProAct 360™ é uma endoprotease com ampla especificidade e clivagem em qualquer ponto da cadeia proteica, o que lhe permite atuar em uma ampla gama de substratos relevantes para a alimentação animal. Ao solubilizar e quebrar a proteína em fragmentos menores, o ProAct 360™ age sinergicamente com proteases endógenas que subsequentemente digerem ainda mais a proteína da ração, permitindo maior absorção pelo animal.
Os testes in vitro indicam a capacidade do ProAct 360™ de aumentar a hidrólise proteica de vários ingredientes importantes da ração, além das proteases digestivas endógenas. Veja a Figura 4.
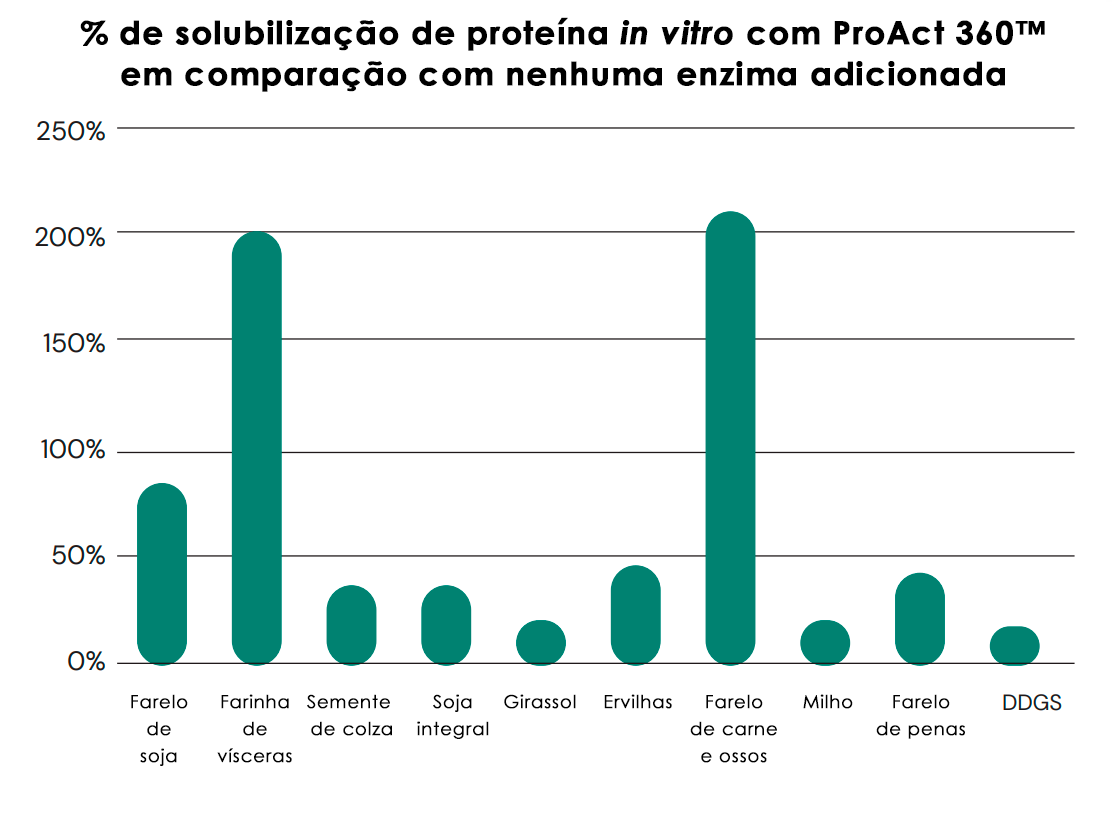 Fig. 4: Solubilização in vitro de diferentes proteínas de matérias-primas relevantes para a alimentação animal. Os substratos e o ProAct 360™ foram incubados por 3 horas a 40°C e pH 5. Após a incubação, o sobrenadante foi analisado pelo ensaio OPA. A solubilização de proteínas com o ProAct 360™ funcionou bem em todas as matérias-primas de teste, aumentando a solubilização de proteínas em 16-84% (A) em substratos vegetais e em 41-211% (B) em relação à amostra controle (sem enzima).
Fig. 4: Solubilização in vitro de diferentes proteínas de matérias-primas relevantes para a alimentação animal. Os substratos e o ProAct 360™ foram incubados por 3 horas a 40°C e pH 5. Após a incubação, o sobrenadante foi analisado pelo ensaio OPA. A solubilização de proteínas com o ProAct 360™ funcionou bem em todas as matérias-primas de teste, aumentando a solubilização de proteínas em 16-84% (A) em substratos vegetais e em 41-211% (B) em relação à amostra controle (sem enzima).
Também é importante sua capacidade de degradar fatores antinutricionais (FAN), por exemplo, do farelo de soja. O farelo de soja contém diferentes fatores antinutricionais, como inibidores de tripsina e lectina, cujo nível depende principalmente das condições de processamento. Diversos estudos indicaram que os níveis de FAN diferem em diferentes lotes de farelo de soja e que o aumento dos níveis de FAN tem consequências negativas no desempenho animal (McNaughton et al. 1981; Palacious et al. 2004; Pacheco et al., 2014). O ProAct 360™ tem o potencial de degradar inibidores de tripsina e lectina no farelo de soja, conforme indicado na Figura 5, ajudando assim a reduzir os efeitos negativos desses FANs no desempenho animal. Também foi demonstrado que os inibidores de tripsina não inibem a ação do ProAct 360™, como a tripsina.
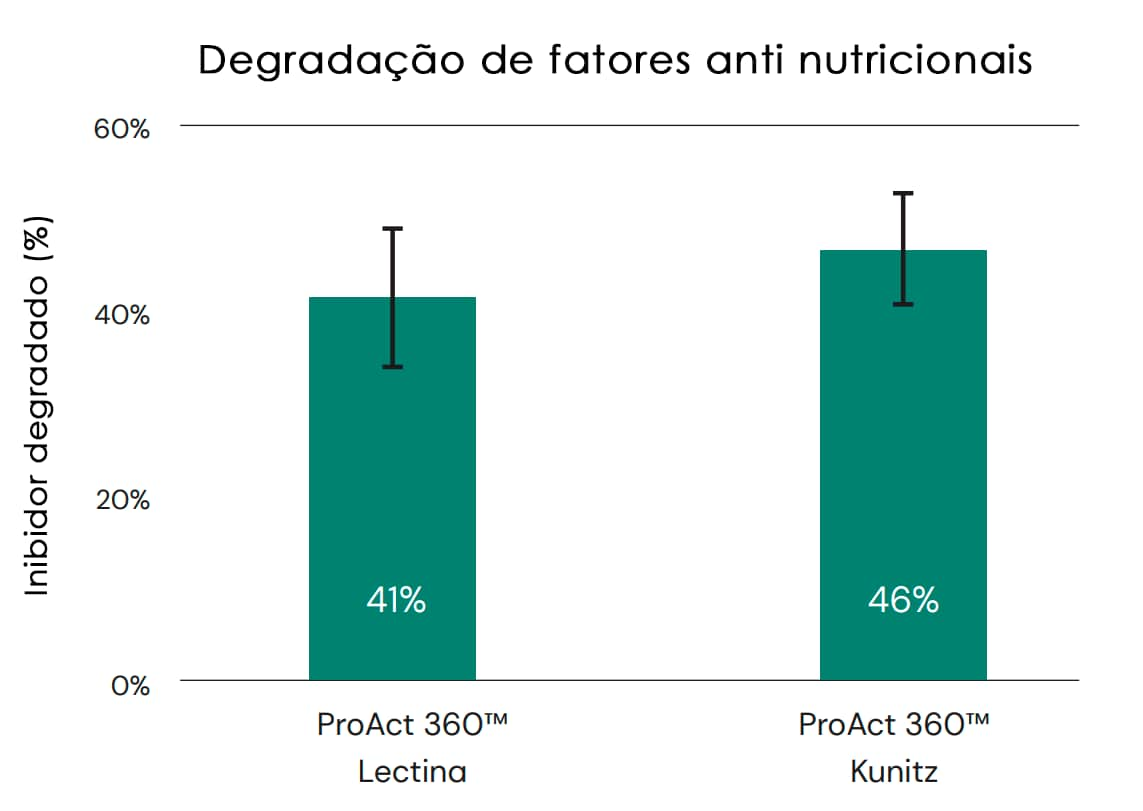 Fig. 5: O ProAct 360™ tem a capacidade de degradar inibidores de tripsina do tipo Lectina e Kunitz do farelo de soja em géis de SDS-Page. As porcentagens indicam a eficiência da degradação calculada a partir da intensidade da banda. A enzima foi misturada com inibidores de tripsina e incubada em pH 7 e 40 °C por 2 horas.
Fig. 5: O ProAct 360™ tem a capacidade de degradar inibidores de tripsina do tipo Lectina e Kunitz do farelo de soja em géis de SDS-Page. As porcentagens indicam a eficiência da degradação calculada a partir da intensidade da banda. A enzima foi misturada com inibidores de tripsina e incubada em pH 7 e 40 °C por 2 horas.
A avaliação in vivo indicou que o perfil de digestibilidade de aminoácidos do ProAct 360™ é semelhante ao da primeira geração do RONOZYME® ProAct. No entanto, foi claramente demonstrado que o ProAct 360™ tinha uma digestibilidade de nitrogênio e aminoácidos consistentemente maior em comparação com o RONOZYME® ProAct (Fig. 6). Essa melhoria pode ser explicada pela maior capacidade do ProAct 360™ de aumentar a hidrólise da proteína, neutralizar o efeito de fatores antinutricionais e complementar as proteases endógenas. A resposta média para o aumento da digestibilidade do nitrogênio no caso do ProAct 360™ foi de +3,2%. Os aminoácidos nutricionalmente relevantes (Met, Cys, Lys, Thr, Trp, Arg, Val, Ile e Leu) variaram de +1,9% para Arg a +5,8% para Cys. Uma melhor digestibilidade do nitrogênio e dos aminoácidos pode resultar em maior ganho de peso e melhor conversão alimentar, além de melhorar o bem-estar animal e a sustentabilidade das empresas avícolas.
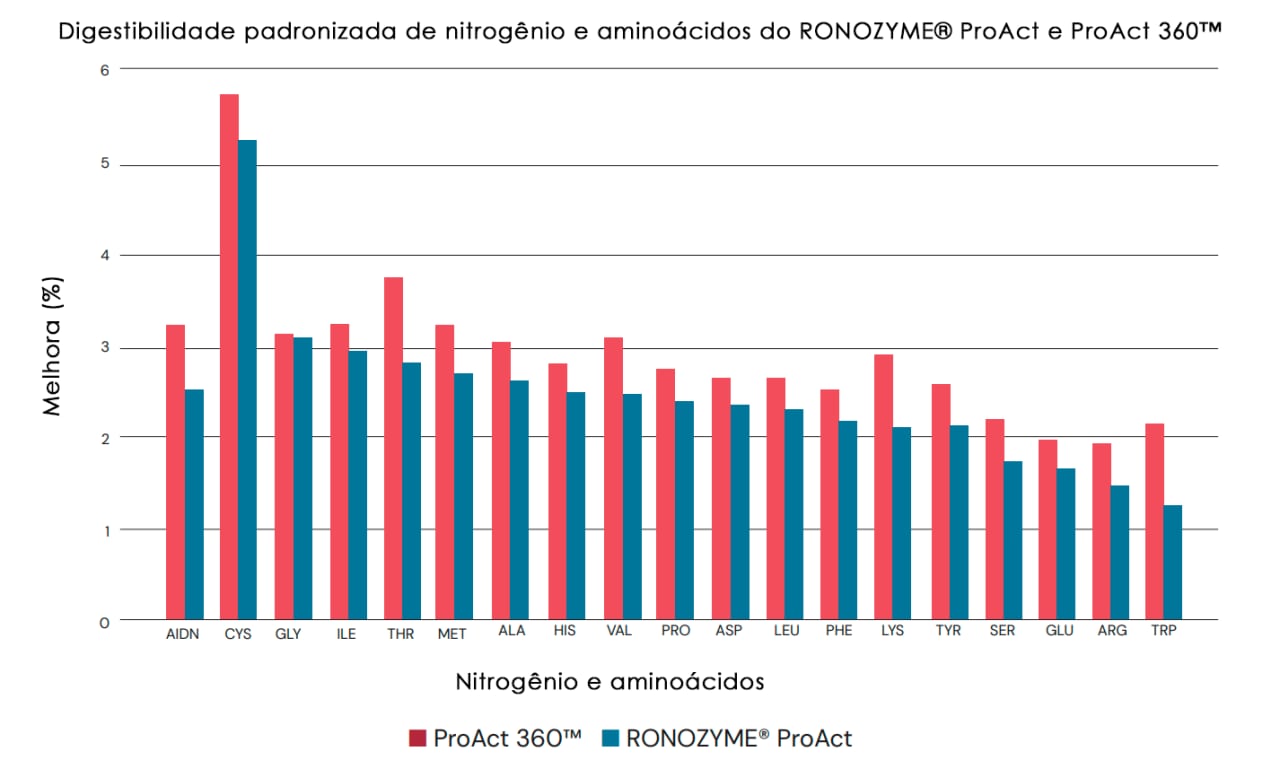 Fig. 6. Perfil de digestibilidade de nitrogênio e aminoácidos do RONOZYME® ProAct (azul) e do ProAct 360™ (vermelho).
Fig. 6. Perfil de digestibilidade de nitrogênio e aminoácidos do RONOZYME® ProAct (azul) e do ProAct 360™ (vermelho).
Além disso, uma protease de alta qualidade deve ser formulada de forma a garantir muitos pequenos grânulos por grama, com homogeneidade de tamanho para assegurar sua miscibilidade na ração. Uma formulação livre de pó também é importante para a segurança do trabalhador. A formulação do ProAct 360™ foi desenvolvida de forma a garantir todos esses parâmetros de segurança e qualidade, a fim de fornecer resultados consistentes e robustos no campo (Fig. 7).
 Fig. 7: Aparência do ProAct 360™. Os grânulos devem ser tão pequenos e homogêneos quanto possível para garantir uma boa miscibilidade na alimentação, com um baixo coeficiente de variação.
Fig. 7: Aparência do ProAct 360™. Os grânulos devem ser tão pequenos e homogêneos quanto possível para garantir uma boa miscibilidade na alimentação, com um baixo coeficiente de variação.
Efeitos diretos
Como o nome sugere, o efeito direto da protease exógena é a hidrólise de polipeptídeos de alto peso molecular em oligopeptídeos de peso molecular mais baixo (Glitsø et al., 2012). Esse efeito depende das características da protease em questão, ou seja, especificidade do substrato, perfil de pH e se é uma protease de ação endógena ou exógena, conforme mencionado acima. O substrato para as proteases exógenas são as proteínas indisponíveis da dieta, os antinutrientes proteicos, como os inibidores de tripsina e as lectinas, bem como as próprias proteínas (em que a taxa de digestão pode aumentar em relação à dependência apenas das proteases endógenas). A hidrólise dessas proteínas aumenta significativamente a digestibilidade das proteínas dietéticas (Cowieson & Roos, 2014), e a magnitude desse aumento depende do aminoácido, da matéria-prima em questão e da qualidade inerente relativa da proteína. É importante observar que há grandes diferenças no efeito da protease por aminoácido e também por matéria-prima, portanto, é necessário prestar atenção à formulação de dietas para acomodar os efeitos da protease. Ou seja, valores fixos de um aminoácido ou matriz proteica podem gerar respostas variáveis em termos de desempenho.
É importante observar que nem todas as proteases exógenas têm uma funcionalidade favorável em dietas para não ruminantes. Cowieson e Roos (2016) resumiram os resultados de uma série de ensaios com proteases exógenas, de origem diversa e mencionadas na literatura científica, e nota-se que muitas delas exercem pouco efeito benéfico sobre as espécies-alvo. Um exemplo concreto disso foi o trabalho de Simbaya et al. (1996), que examinaram a eficácia de uma série de cinco proteases exógenas diferentes para frangos de corte. Eles descobriram que apenas uma dessas proteases teve um efeito benéfico sobre o desempenho dos pintinhos e, mesmo assim, a extensão do efeito dependeu do teor de proteína e da fonte da dieta à qual foi adicionada. Recentemente, Walk et al. (2018) publicaram resultados semelhantes sobre a incapacidade de demonstrar a eficácia de novas proteases experimentais. Isso estaria associado ao acesso a uma diversidade limitada de candidatos adequados a proteases, à formulação excessivamente simplista da dieta (sem levar em conta os efeitos diferenciais entre aminoácidos e matérias-primas) e ao não reconhecimento de possíveis diferenças de efeito de acordo com a espécie e a idade do animal.
Efeitos indiretos
Embora o efeito direto da protease exógena seja a hidrólise do substrato - antinutrientes proteicos e proteínas da dieta que, de outra forma, não seriam digeridos no intestino - há vários efeitos "extraproteicos" significativos (não muito diferentes dos efeitos extrafosfóricos da fitase). Esses efeitos estão associados aos benefícios da protease na saúde intestinal (Peek et al., 2009; Kalmendahl e Tauson, 2012; Cowieson et al., 2015) e à melhor digestibilidade de nutrientes alternativos não proteicos, como a energia (mediada pelo aumento da digestibilidade da gordura e do amido).
Uma preocupação comum quando se trata do uso de várias enzimas na mesma dieta é a canibalização do valor. Não há dúvida de que, embora as diferentes classes de enzimas (fitases, carboidrases e proteases) não compartilhem os mesmos substratos, a hidrólise desses diferentes substratos gera vários efeitos semelhantes na digestibilidade, que têm um efeito cumulativo (embora provavelmente não totalmente aditivo) no desempenho animal. Por exemplo, foi demonstrado que as carboidrases (principalmente xilanases e glucanases) (Cowieson, 2010); fitase (Cowieson et al., 2018b,c); e protease (Cowieson e Roos, 2014) aumentam a digestibilidade ileal de aminoácidos em animais não ruminantes. No caso da melhor digestibilidade de aminoácidos com enzimas exógenas, isso se deve tanto a uma redução no fluxo de aminoácidos endógenos (como é principalmente o caso da fitase) quanto a uma verdadeira melhoria na digestibilidade da proteína dietética (como é o caso das proteases e carboidrases). A falta de aditividade entre essas classes de enzimas está ligada principalmente ao efeito de sobreposição na perda endógena (como é bem observado na avaliação da digestibilidade de aminoácidos em ingredientes individuais, o que levou ao desenvolvimento do ensaio DIE). No caso da fitase, os efeitos sobre a digestibilidade de aminoácidos podem estar diretamente ligados à redução da perda de proteína endógena (principalmente mucina, mas também pepsina). Esses efeitos estão relacionados ao efeito do fitato na solubilidade da proteína no intestino e ao aumento resultante na produção endógena de mucina e pepsina (resumo de Cowieson et al., 2009).
De fato, Cowieson et al. (2008) observaram uma correlação direta entre o efeito do fitato e da fitase no fluxo de aminoácidos endógenos em frangos de corte e o perfil de aminoácidos da mucina e da pepsina. Além disso, a estimulação da perda excessiva de aminoácidos endógenos pela ingestão de ácido fítico pode ser facilmente resolvida pela inclusão de concentrações relativamente baixas de fitase microbiana, sem outros benefícios observados quando a inclusão aumenta acima de aproximadamente 500-750 FYT/kg (Cowieson et al., 2018 b,c). Esses resultados foram observados anteriormente (Shirley & Edwards, 2003; Fig. 8). Esse fato sugere que qualquer canibalização do valor das carboidrases ou proteases pela fitase na digestibilidade dos aminoácidos será evidente em baixas concentrações de inclusão de fitase, e isso não se altera à medida que a dose de fitase é aumentada.
No caso das carboidrases, os efeitos sobre a digestibilidade dos aminoácidos parecem não estar fortemente ligados ao fluxo endógeno de aminoácidos.
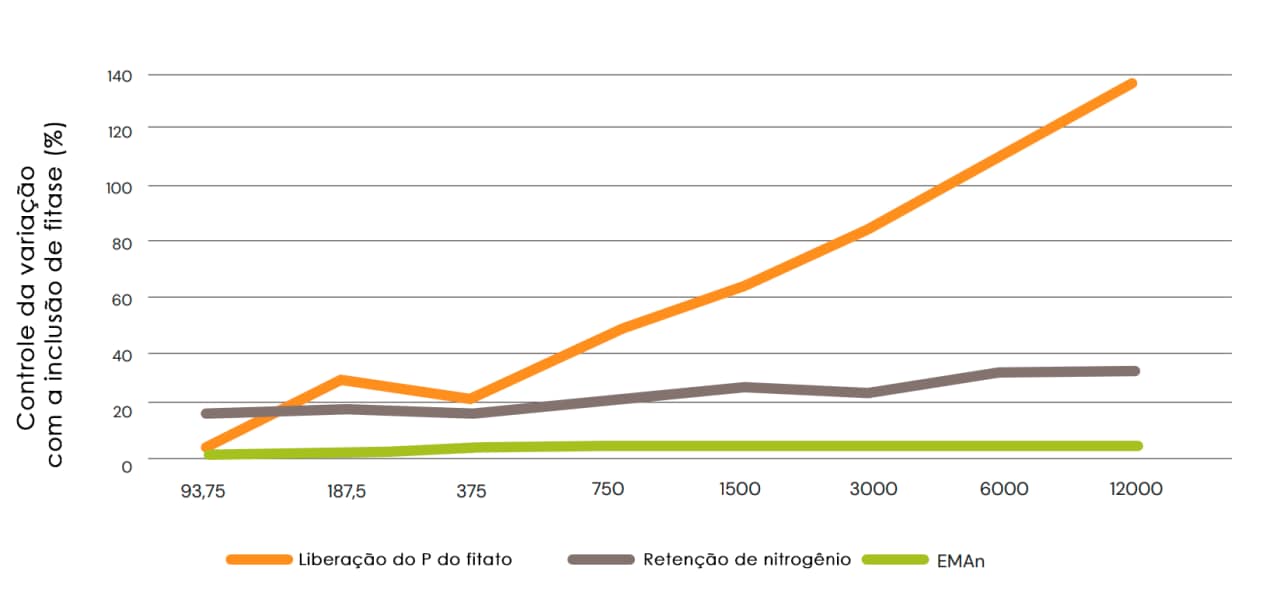 Fig. 8: Efeito do aumento da dose de fitase na liberação do P do fitato, na retenção de nitrogênio e na energia metabolizável aparente corrigida pelo nitrogênio (EMAn) em frangos de corte alimentados com dietas de milho/farelo de soja (Shirley & Edwards, 2003).
Fig. 8: Efeito do aumento da dose de fitase na liberação do P do fitato, na retenção de nitrogênio e na energia metabolizável aparente corrigida pelo nitrogênio (EMAn) em frangos de corte alimentados com dietas de milho/farelo de soja (Shirley & Edwards, 2003).
Isso se baseia em uma grande meta-análise que mostra que nenhum aminoácido específico se beneficia mais do que outro com a adição de xilanase ou glucanase, o que não seria o caso se o fluxo de proteína endógena fosse desproporcionalmente influenciado em relação à digestibilidade da proteína da dieta (Cowieson e Bedford, 2009). Isso sugere que o efeito das carboidrases sobre a digestibilidade ileal dos aminoácidos pode ser amplamente aditivo ao efeito da fitase, já que a primeira modifica a digestibilidade da dieta (presumivelmente por meio de mecanismos genéricos que envolvem a permanência gástrica da ração, a fermentação do intestino grosso, a redução da viscosidade do conteúdo do lúmen e a fuga do conteúdo da parede celular), enquanto a última modifica o fluxo de proteína endógena.
Por fim, as proteases proporcionam melhorias na digestibilidade de aminoácidos, por um lado, reduzindo a perda de mucina endógena (aproximadamente 30% de seu efeito) e, por outro lado, aumentando a retenção de aminoácidos da dieta (Cowieson & Roos, 2016). Portanto, pode-se esperar alguma diminuição no efeito da fitase sobre a digestibilidade de aminoácidos com a adição de protease, especialmente para aminoácidos comumente encontrados na proteína endógena, por exemplo, Thr, Cys, Gly, Ser e Pro.
A soja armazena proteínas em vacúolos de armazenamento de proteínas (VAPs), que são estruturas globulares compostas principalmente de proteínas, ácido fítico e minerais (Jiang et al. 2001). O ácido fítico está concentrado em partes densas dos VAPs chamadas globóides de fitato. Como o fitato presente nesses globoides pode ser incorporado às proteínas, não se espera que ele esteja prontamente acessível para a desfosforilação por uma fitase. Bohn et al. (2007) demonstraram que a fitase degrada o fitato livre e solúvel muito mais rapidamente do que o fitato presente nos globoides de fitato. As proteínas globóides podem retardar a ação da fitase ao bloquear o acesso ao ácido fítico. Foi demonstrado que o ProAct 360™ degrada as proteínas presentes nos vacúolos de armazenamento de proteínas (VAPs), levando a uma maior solubilização do ácido fítico, que pode então ser facilmente desfosforilado pela fitase.
O efeito do ProAct 360™ nos vacúolos de armazenamento de soja (VAPs) é mostrado na Figura 9. Seções de tecido de cotilédone de soja foram fixadas, desidratadas e incubadas com ProAct 360™, e o efeito da protease foi visualizado por microscopia eletrônica de varredura. Foram observados inúmeros orifícios nos vacúolos de armazenamento de proteína do tecido tratado com enzima, demonstrando a degradação da proteína presente nos VAPs.
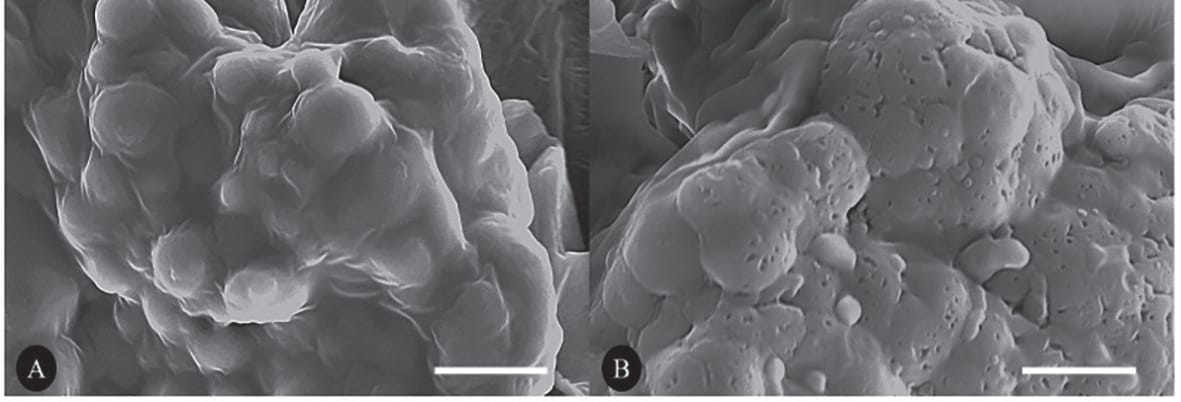 Fig. 9: Seções de tecido fixo de cotilédone de soja não tratadas/tratadas com ProAct360™ e visualizadas por microscopia eletrônica de varredura. Numerosos orifícios são observados nos vacúolos de armazenamento de proteína do tecido tratado com enzima. (A) Controle sem protease; (B) com adição de protease.
Fig. 9: Seções de tecido fixo de cotilédone de soja não tratadas/tratadas com ProAct360™ e visualizadas por microscopia eletrônica de varredura. Numerosos orifícios são observados nos vacúolos de armazenamento de proteína do tecido tratado com enzima. (A) Controle sem protease; (B) com adição de protease.
Considerar as diferenças nas matérias-primas
Não há dúvida de que as proteases exógenas não influenciam todas as fontes de proteína na mesma medida (Figura 10). Isso provavelmente está relacionado à especificidade de uma determinada protease para diferentes tipos de proteínas, por exemplo, proteínas hidrofóbicas ou polares, cargas variáveis, sequências de aminoácidos, etc., bem como à digestibilidade inerente dos aminoácidos de diferentes matérias-primas.
Esse também é o caso do pool de proteases endógenas, em que algumas proteínas da dieta demonstraram escapar amplamente da digestão. Por exemplo, Cowieson et al. (2017) usaram a proteômica para determinar a origem dos peptídeos no íleo de frangos de corte alimentados com uma dieta à base de milho/soja e observaram que a maioria dos fragmentos de peptídeos era proveniente de proteínas de armazenamento não digeridas do farelo de soja. Portanto, não é surpreendente que proteases exógenas específicas também "favoreçam" algumas fontes de proteína mais do que outras e, portanto, apresentem compatibilidade variável com diferentes ingredientes de ração. O efeito da protease in vivo pode ser otimizado por meio de uma avaliação cuidadosa da mistura da dieta (fonte e proporcionalidade da proteína da dieta) para desenvolver uma matriz de resposta apropriada para essa dieta específica.
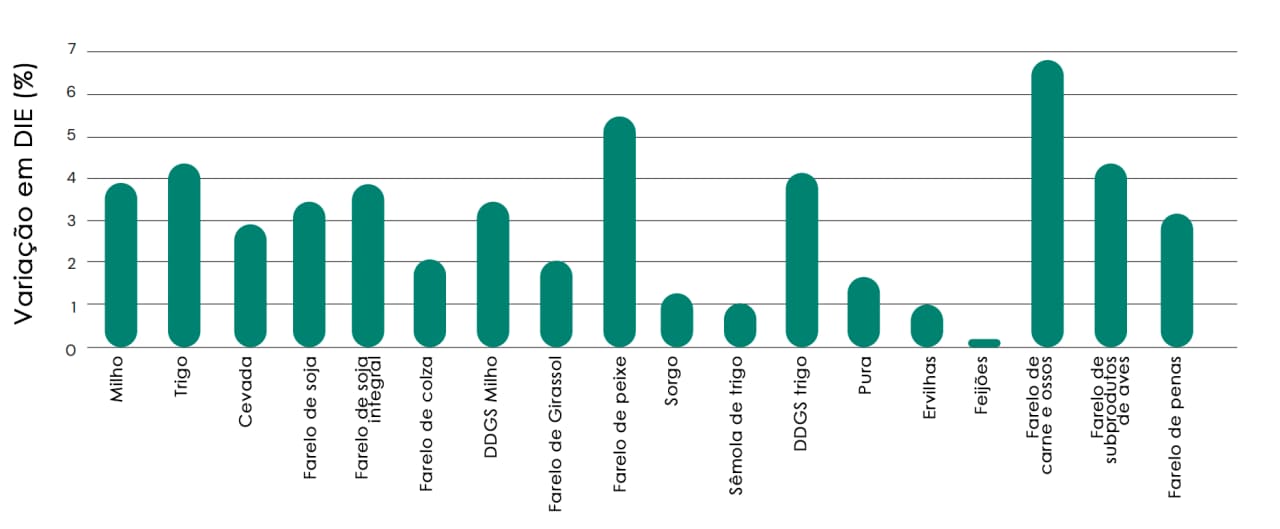 Fig. 10: Efeito do ProAct 360™ na digestibilidade ileal estandardizada de aminoácidos (DIE; média de 9 aminoácidos) em ingredientes comuns de ração. DDGS de milho = grãos de destilação de milho com solúveis; DDGS de trigo = grãos de destilação de trigo com solúveis.
Fig. 10: Efeito do ProAct 360™ na digestibilidade ileal estandardizada de aminoácidos (DIE; média de 9 aminoácidos) em ingredientes comuns de ração. DDGS de milho = grãos de destilação de milho com solúveis; DDGS de trigo = grãos de destilação de trigo com solúveis.
Considerar a divergência entre os aminoácidos
A formulação de dietas para animais não ruminantes depende em grande parte da manutenção da densidade adequada de aminoácidos e também das proporções de lisina para garantir um suprimento equilibrado de aminoácidos para apoiar o crescimento. Como as proteases exógenas não liberam aminoácidos na mesma proporção (Figura 6), para que os efeitos benéficos sobre a digestibilidade resultem em ganhos de desempenho mensuráveis, é fundamental que a formulação da protease seja desenvolvida reconhecendo as respostas divergentes dos aminoácidos. Essas respostas são específicas para ingredientes individuais da ração, mas também são específicas para aminoácidos específicos. Em geral, no que se refere à lisina (Lys), as melhorias associadas à protease são mais substanciais para Thr, Cys, Gly, Pro, Ala e Val e menos substanciais para Glu, Trp, Phe, His, Ile, Met, Tyr, Leu, Arg, Asp e Ser (Fig. 11). Isso significa que o uso de protease pode alterar as proporções ideais de aminoácidos em uma determinada dieta. E, de fato, isso acontecerá se for aplicada uma matriz plana de protease, que não reconhece as diferentes magnitudes de resposta por aminoácido. Portanto, é essencial que a formulação de dietas que envolvam o uso de proteases não seja feita de forma simplista ou sem considerar a influência de uma combinação específica de matérias-primas, além da densidade e do equilíbrio de aminoácidos.
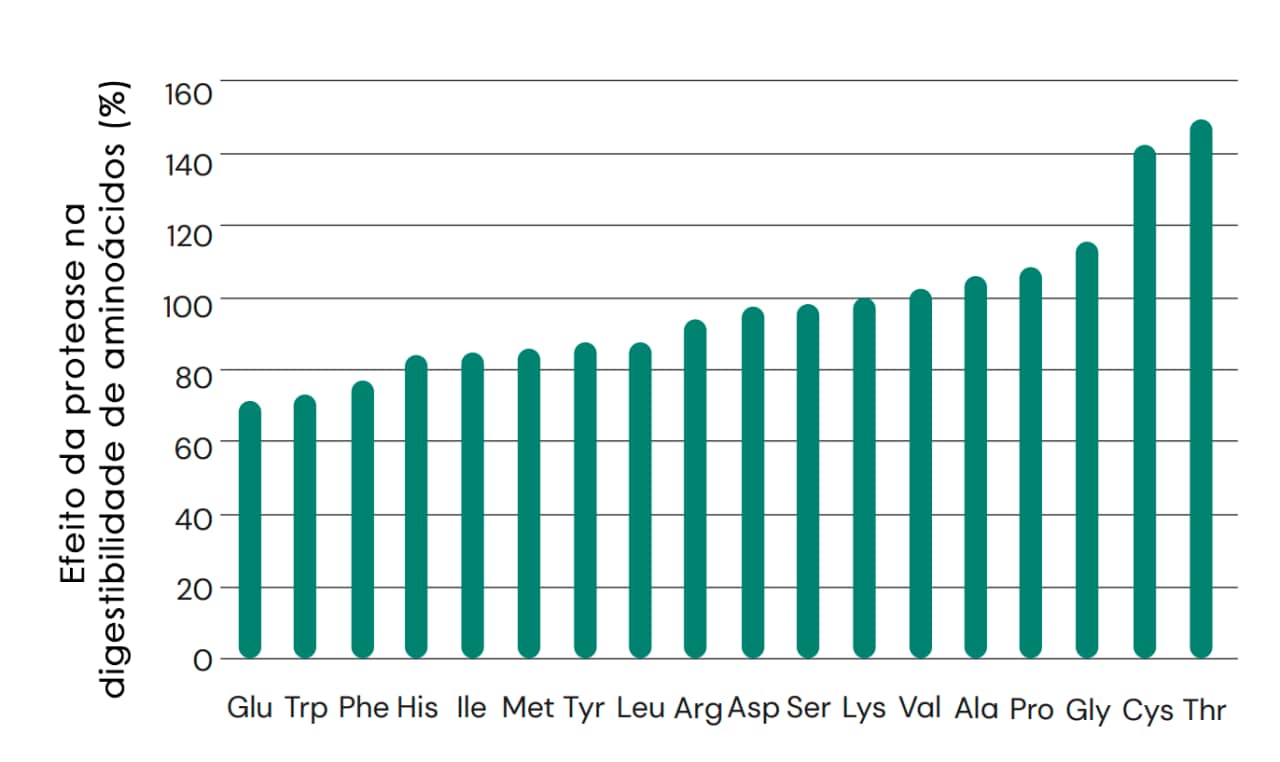 Fig. 11: Efeito do ProAct 360™ na digestibilidade ileal de aminoácidos em animais não ruminantes, expresso em relação à lisina (Lys) (Cowieson e Roos, 2014).
Fig. 11: Efeito do ProAct 360™ na digestibilidade ileal de aminoácidos em animais não ruminantes, expresso em relação à lisina (Lys) (Cowieson e Roos, 2014).
Factores adjacentes
Além do efeito da protease exógena em matérias-primas e aminoácidos específicos, há várias outras considerações que podem influenciar a magnitude e a consistência do efeito (para uma análise detalhada, consulte Cowieson e Roos, 2016). Um desses fatores é a qualidade da proteína em uma determinada matéria-prima. Por exemplo, um lote de farelo de soja com digestibilidade inerentemente baixa apresentará uma resposta elevada à protease em comparação com um lote de qualidade superior. Foi demonstrado de forma conclusiva que os efeitos da protease são substancialmente maiores e mais consistentes quando a digestibilidade ileal dos aminoácidos em uma dieta ou matéria-prima específica tem valores abaixo de 90% em comparação com valores mais altos (Cowieson e Roos, 2014), o que oferece uma margem considerável para intervenção estratégica (Figura 12). Por exemplo, fatores capazes de promover ou degradar a digestibilidade da proteína podem estar conceitualmente ligados ao uso de protease exógena, como espécie animal, idade, processamento da ração, tamanho da partícula etc. A esse respeito, é importante observar que o desenvolvimento de uma dieta de "controle negativo", para examinar o efeito da protease exógena por meio da remoção da proteína da ração, provavelmente será desvantajoso, pois: a) o substrato é removido; b) o equilíbrio de aminoácidos é perdido; c) o animal reage à insuficiência proteica por meio da regulação positiva dos transportadores de peptídeos, o que aumenta a digestibilidade da proteína restante na dieta. Além disso, é essencial assegurar um suprimento adequado de nutrientes limitantes, como energia e fósforo, para garantir que o efeito benéfico da protease não seja perdido.
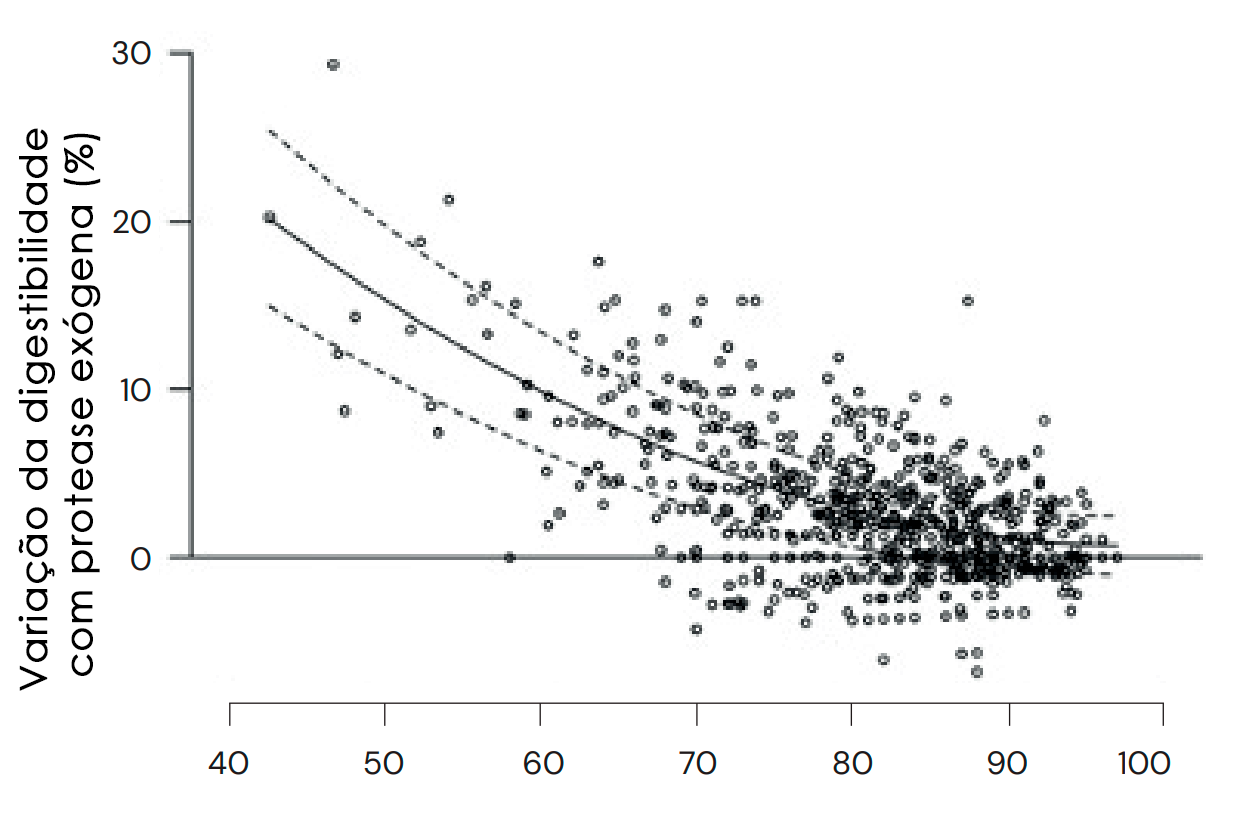 Fig. 12: Os efeitos do ProAct360™ são substancialmente maiores e mais consistentes quando a digestibilidade ileal dos aminoácidos em uma dieta/matéria-prima específica é inferior a 90%.
Fig. 12: Os efeitos do ProAct360™ são substancialmente maiores e mais consistentes quando a digestibilidade ileal dos aminoácidos em uma dieta/matéria-prima específica é inferior a 90%.
O desenvolvimento de proteases exógenas que sejam funcionais em todos os animais monogástricos, suínos, aves e espécies aquáticas é extremamente difícil. Muitas dessas enzimas não funcionam, ou não funcionam bem, e não são compatíveis com as enzimas endógenas ou com o tipo de substrato presente nos ingredientes comuns das rações. Além disso, uma vez identificada uma protease adequada, não se pode simplesmente adicioná-la a uma dieta e observar diretamente as respostas úteis de desempenho. Em vez disso, é necessário um trabalho considerável para mapear o efeito da protease em várias matérias-primas e aminoácidos para garantir a formulação estratégica da dieta. Também é importante observar que o efeito benéfico das proteases exógenas não se limita à melhor digestibilidade dos aminoácidos, mas também se estende à distribuição/eficiência energética, à sustentabilidade ambiental, à saúde intestinal e a outros parâmetros importantes de produção.
Por fim, embora seja possível esperar uma pequena redução no benefício dos aminoácidos quando várias enzimas são adicionadas à mesma dieta, é provável que esses efeitos sejam modestos e especificamente direcionados aos aminoácidos encontrados na proteína endógena, por exemplo, treonina.
Portanto, pode-se concluir que as proteases exógenas são eficazes para melhorar a sustentabilidade dos sistemas de produção de animais não ruminantes. No entanto, as proteases exógenas não podem ser usadas de forma simplista, sem primeiro ter o conhecimento básico para usá-las de forma eficaz. Uma vez que essa base tenha sido estabelecida, as proteases exógenas oferecem um potencial significativo para melhorar a lucratividade da avicultura e da suinocultura em todo o mundo, bem como para manter a saúde e o bem-estar dos animais, diante dos desafios significativos associados à alimentação de uma população mundial em crescimento.
2 abril 2024
We detected that you are visitng this page from United States. Therefore we are redirecting you to the localized version.
