


-
English
- Global
- North America
- Latin America
La glucosa es clave para el metabolismo de la energía en las aves y porcinos y se transforma eficientemente en ATP por medio del Ciclo del Acido Cítrico, para su utilización en diversos procesos que requieren energía. Gran parte de la glucosa que ingieren las aves y los porcinos procede del almidón, generalmente obtenida de diversos cereales. El almidón es un polímero de glucosa que consiste de enlaces α-1,4 y/o α-1,6. Cuando se produce un enlace α-1,4 entre los monómeros de glucosa, el polímero resultante se denomina amilosa, mientras que cuando se produce un enlace α-1,6, el polímero resultante se conoce como amilopectina. Debido a la orientación de los monómeros de glucosa adyacente en los polímeros de amilasa, se desarrolla una hélice natural con un exterior hidrofílico y un núcleo hidrofóbico. Estas hélices de amilosa rápidamente forman cristales moderadamente insolubles y bastante inaccesibles para la digestión de las enzimas. La amilopectina -por otra parte- tiene una macro-estructura más desordenada dado que los puntos de ramificación α-1,6 interrumpen la estructura de la hélice creando regiones amorfas a las que a menudo se hace referencia como "cerosas". Por lo tanto, las regiones altamente ramificadas de almidón ceroso dentro del endospermo de los cereales es a menudo un punto de entrada para que las amilasas pancreática y de la saliva inicien la degradación del almidón a oligómeros de dextrina. La relación entre la amilosa y la amilopectina en el almidón determina si un almidón se puede categorizar como "ceroso" o "altamente amiloso" y esto varía entre los distintos tipos de granos de cereales. Un maíz ceroso, por ejemplo, puede contener hasta un 99% de amilopectina mientras que un maíz con alto contenido de amilosa puede contener más de un 70% de amilosa. Asimismo, hay una heterogeneidad considerable en el tamaño del gránulo de almidón en semillas provenientes de diversas plantas y en la naturaleza de micro y macro estructuras cristalinas que favorecen -o no- la digestión. Siguiendo la hidrólisis inicial de las amilasas endógenas, el proceso de la digestión del almidón se completa con las enzimas maltasa e iso-maltasa, y la glucosa resultante se absorbe por medio de un sistema de transporte dependiente del Na, para su metabolismo eventual a través del Ciclo del Acido Cítrico. Dado que la mayoría de las dietas para aves y porcinos contienen 400-500 g/kg de almidón, la conversión exitosa del almidón en glucosa es extremadamente importante para el estado energético del animal y esto se comprueba en los animales de más edad con mayor consumo de almidón. Por lo tanto, los factores que demoran, dificultan o interfieren con la solubilización y digestión del almidón tienen un profundo impacto sobre el valor energético de cada dieta. Dichos factores incluyen la naturaleza de los cristales del almidón, la extensión del daño que sufre el almidón durante el procesamiento hidrotérmico del alimento o los ingredientes, factores que pueden inhibir el óptimo funcionamiento de las amilasas endógenas y cuestiones relacionadas con la extracción de glucosa del lúmen a partir de sistemas de transporte dependientes del Na.
El propósito de este breve artículo es tratar resumidamente dichos factores y delinear la utilidad de las fuentes exógenas de las amilasas en la nutrición de animales monogástricos, con énfasis tanto en los efectos directos de la energía digerible como así también en los efectos indirectos de la energía neta y los requerimientos de amino ácidos.
El almidón tiene una estructura extremadamente heterogénea que varía en la composición entre las amilosas y amilopectinas, en solubilidad, cristalinidad e interacción con otros micro y macro nutrientes (Moran, 1982, Tester et al., 2004) Los cristales característicos del almidón se encuentran generalmente ya sea como gránulos tipo A (se encuentran más comúnmente en los cereales); gránulos tipo B (en los tubérculos) o tipo C (una mezcla de los tipos A y B, que se encuentra a menudo en las legumbres) (Tester et al., 2004).
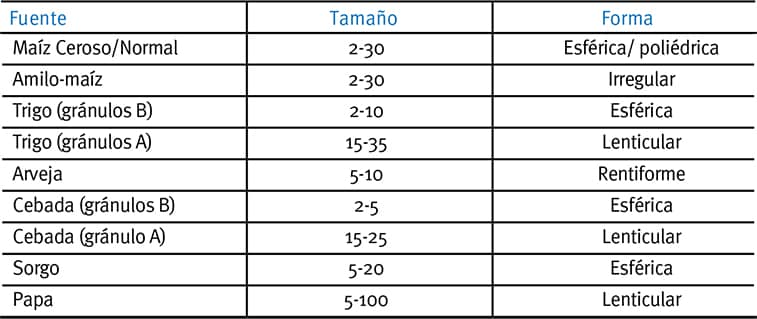 Tabla 1. Características de los gránulos de almidón nativo procedentes de diversos cereales, legumbres y tubérculos (recomposición de Tester et al., 2004).
Tabla 1. Características de los gránulos de almidón nativo procedentes de diversos cereales, legumbres y tubérculos (recomposición de Tester et al., 2004).
 Figura 1: El gránulo de almidón visto con microscopía electrónica de barrido (izquierda a derecha) de papas, arroz y trigo (con autorización de Prof. E.T. Moran).
Figura 1: El gránulo de almidón visto con microscopía electrónica de barrido (izquierda a derecha) de papas, arroz y trigo (con autorización de Prof. E.T. Moran).
Los gránulos tipo A son más compactos y contienen menos agua que los gránulos tipo B (Wu & Sarko, 1978a, b). Tester et al. (2004) señalaron la característica del gránulo de almidón nativo en diversos cereales, legumbres y tubérculos (extracto en la Tabla 1 y en la Figura 1) revelando la heterogeneidad del tamaño y forma de los granos que existen en los ingredientes nutricionales más comunes para cerdos y aves.
El almidón se define como ceroso, normal o "amilo" cuando el rango de amilosa a amilopectina está cerca del 15%, 16-35% o >36% respectivamente (Tester et al., 2004; Tabla 2; Fig. 2). Debido a los efectos de las ramas de amilopectinas de los enlaces α-1,6 sobre las formaciones de cristales, los almidones con alta proporción de amilopectina relativos a la amilosa tienden a ser más amorfos y más propensos a la digestión que los amilo-almidones.
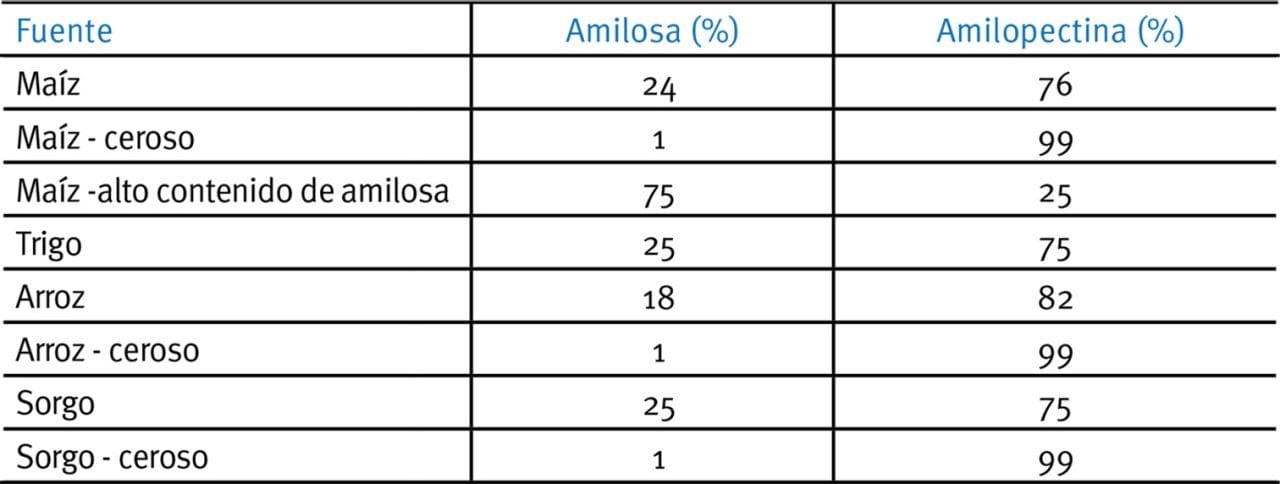 Tabla 2. Relación de amilosa / amilopectina en distintos granos (con autorización del Prof. E.T. Moran).
Tabla 2. Relación de amilosa / amilopectina en distintos granos (con autorización del Prof. E.T. Moran).
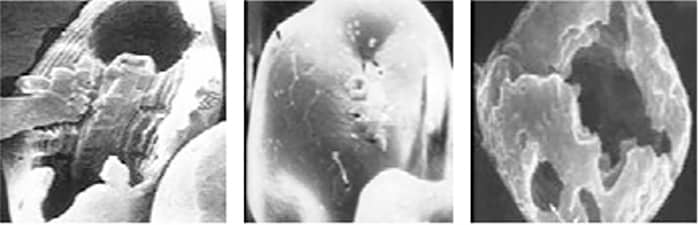 Fig. 2: Corn starch granules under electron microscopy: from left to right (amylopectin:amylose), normal (75:25), high amylose (25:75) or waxy (99:1). (Used with permission from Prof. E.T. Moran.)
Fig. 2: Corn starch granules under electron microscopy: from left to right (amylopectin:amylose), normal (75:25), high amylose (25:75) or waxy (99:1). (Used with permission from Prof. E.T. Moran.)
Adicionalmente, en los almidones de los cereales únicamente los lípidos están asociados a los residuos de amilosa y aproximadamente el 1.5% de la masa de gránulos pueden ser lípidos, especialmente en almidones de alto contenido de amilosa (Morris, 1993).
El maíz es la fuente de almidón dominante en la dieta global de los porcinos y aves (Cowie, 2005) y ofrece el mayor aporte de energía para la dieta. El almidón de maíz se encuentra predominantemente en el endospermo (aprox. 86%), aunque un 4-5% del almidón se encuentra en el germen y en las capas de aleurona, respectivamente. Es importante destacar que en el maíz, los gránulos de almidón se encuentran dentro de una matriz de proteínas hidrofóbica y que puede estar encastrada de forma compacta o suelta, dependiendo de las condiciones genéticas o de crecimiento.
Cuando la matriz de almidón/proteínas está encastrada de forma compacta y es alta en proteínas en relación con el almidón, el endospermo se describe como flint o altamente vidrioso y cuando la matriz está encastrada de manera floja y se puede desmenuzar, el endospermo se describe como harinoso (Figura 3).
Por ello, el almidón no debería considerarse una fracción independiente y homogénea, sino un compuesto complejo heterogéneo que consiste de partes variables de cristales y zonas amorfas, ípidos adherentes y, a menudo, capas de aleurones de hemicelulosas. La digestión exitosa del almidón requiere la solubilización de amilosas hidrofóbicas, disociaciones de macro nutrientes adheridos y una producción adecuada de enzimas endógenas relacionadas con la capacidad de absorción y el volumen del alimento ingerido.
 Fig. 3: Milho altamente (A) e pouco (B) vítreo (Gibson et al., 2003), mostrando os grânulos de amido incorporados a uma matriz de proteína prolamina no endosperma.
Fig. 3: Milho altamente (A) e pouco (B) vítreo (Gibson et al., 2003), mostrando os grânulos de amido incorporados a uma matriz de proteína prolamina no endosperma.
La digestión del almidón en porcinos se inicia en la boca por acción de la amilasa de la saliva mientras que en las aves no se produce hasta que la amilasa pancreática entra en contacto con los polímeros del almidón ingerido (Moran, 1988) La α-amilasa pancreática hidroliza amilosa en maltosa y maltotriosa y la amilopectina, en malteosa, maltotriosa y dextrinas α-límite; estos oligosacáridos luego se degradan a glucosa - en la capa de agua superficial por la maltasa-, y en sucrosa isomaltasa -en la superficie de las micro-vellosidades (Morán, 1985). La glucosa resultante se transporta por medio del sistema de transporte dependiente del Na hacia el citoplasma de la célula y a lo largo de la membrana baso-lateral para un eventual ingreso al Ciclo del Acido Cítrico para generación de ATP. En las aves, el íleon terminal digiere casi completamente (a menudo >95%) el almidón, pero esta situación varía y depende de la edad, la estructura y solubilidad del almidón y el procesamiento hidrotérmico de las dietas y otros factores diversos.
Aparentemente, las aves desarrollan un aumento en la capacidad para digerir almidón dado que su tracto intestinal madura y se evidencia una elevada producción de amilasa pancreática en aves de mayor edad en comparación con aves más jóvenes (Krogdahl & Sell, 1989). Asimismo, las aves de corral desarrollan la capacidad de digerir el almidón en el huevo (in ovo) (Moran, 1985) y después de la eclosión, tienen una mayor capacidad para digerir el almidón en comparación con los mamíferos, como por ejemplo los lechones que dependen de la lactosa como fuente de energía dominante. El rápido incremento de la producción de amilasa pancreática en aves jóvenes ha generado especulaciones con respecto a que este aumento podría ser una respuesta por la utilización de amilasas microbianas exógenas.
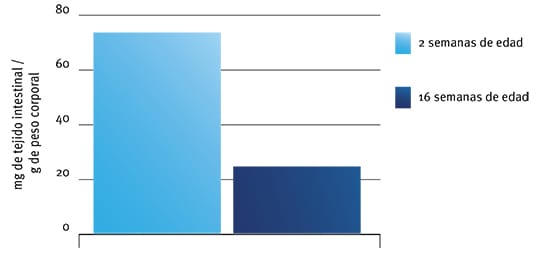 Figura 4: Efectos de la edad sobre el peso relativo del intestino en pavos (a partir del trabajo de Croom et al., 1999).
Figura 4: Efectos de la edad sobre el peso relativo del intestino en pavos (a partir del trabajo de Croom et al., 1999).
Sin embargo, cuando se considera el consumo de almidón (especialmente por unidad de tejido intestinal) son realmente los animales de mayor edad los que podría beneficiarse más por el suministro de amilasas exógenas”. Por ejemplo, Croom et al. (1999) observaron que un pavo de 2 semanas de edad tenía casi cuatro veces más tejido intestinal por gramo de peso corporal que un pavo de 16 semanas (Figura 4).
Estos datos, que también se aplican a pollos de engorde, sugieren que el intestino tiene una importancia notable en las aves jóvenes y su desarrollo exitoso temprano es extremadamente importante para asegurar el mantenimiento adecuado de las aves en las etapas de crecimiento posteriores. Asimismo, Lilja (1980, 1983) sugirió que el desarrollo exitoso (especialmente en aves de un alto índice de crecimiento) en las últimas etapas de crecimiento depende del desarrollo adecuado de los intestinos del recién nacido; esta conclusión fue respaldada por Pinchasov et al., (1985).
 Figura 5: Producción de las enzimas digestivas en los tejidos pancreáticos o intestinales de las aves en función de los días de edad (expresado en un porcentaje de valores respecto a la semana 8). (Krogdhal & Sell (1989).
Figura 5: Producción de las enzimas digestivas en los tejidos pancreáticos o intestinales de las aves en función de los días de edad (expresado en un porcentaje de valores respecto a la semana 8). (Krogdhal & Sell (1989).
Por ello, a pesar de que Krogdhal & Sell (1989; Fig. 2) y Uni et al. (1998) demostraron claramente que la función intestinal en los recién nacidos no está totalmente madura hasta los 14 días post-eclosión, el grado de insuficiencia puede no resultar evidente hasta que se generan altas demandas de nutrientes en los intestinos en aves de peso corporal elevado, desde el día 35 hasta la faena. En este sentido, el desarrollo de intestinos y esqueleto tienen consecuencias similares, siendo fundamental el desarrollo temprano, aunque las consecuencias de estas insuficiencias pueden no evidenciarse hasta varias semanas posteriores a la eclosión. Por ello, a pesar de que la actividad enzimática per se (Fig. 5) aumenta con la edad en términos de unidades de actividad por gramo de tejido de páncreas o intestino, la cantidad absoluta de enzimas digestivas por unidad de peso corporal pueden no aumentar. Por ejemplo, Krogdahl & Sell (1989) demostraron que el peso relativo del páncreas aumenta desde un 0.1% a 0.5%, del día 1 al día 10 y luego disminuye hasta un 0.15% hacia el día 56 (Figura 6). Estos resultados concuerdan con lo observado por Croom.
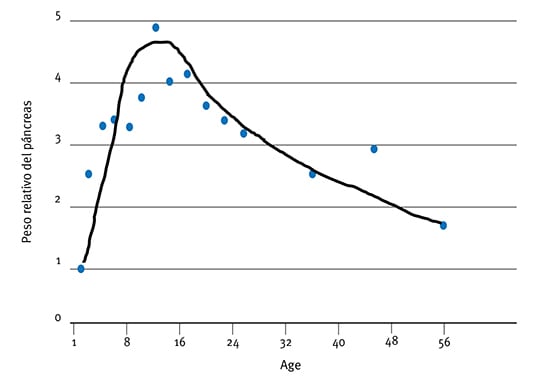 Figura 6: Desarrollo del peso relativo del páncreas en pavos, como porcentaje de peso vivo.
Figura 6: Desarrollo del peso relativo del páncreas en pavos, como porcentaje de peso vivo.
La importancia de estos cambios en el desarrollo de los intestinos relativos al aumento del peso corporal, puede ser contra-intuitivo para el uso de enzimas exógenas, en el sentido que mientras las aves jóvenes tienen claramente una capacidad limitada para producir enzimas endógenas, el tracto intestinal ofrece una proporción mayor de su masa corporal que en pollos de engorde en etapas de crecimiento y terminación. Por eso, podría ser más apropiada la suplementación con enzimas como la amilasa en pollos de engorde pesados donde los tejidos intestinal y pancreático disminuyen proporcionalmente al peso metabólico del ave. Croom et al., (1999) y Krogdahl & Sell (1989), demostraron que a pesar de que la actividad enzimática del intestino al día 8 está cerca del 40% de la actividad del día 56 por unidad de tejido intestinal, como porcentaje del peso corporal existe aproximadamente 4 veces más tejido intestinal en un ave joven que en un ave de mayor edad. Estos resultados sugieren que la masa intestinal y la actividad enzimática pueden ser más limitantes en las aves a partir del día 35 que en las aves más jóvenes, o al menos que el criterio que sólo las aves jóvenes necesitan enzimas exógenas para aumentar la función intestinal puede no ser enteramente correcta.
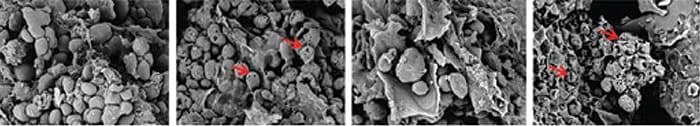 Figura 7: Microscopía electrónica de barrido de una dieta de etapa iniciación de pollos de engorde basada en maíz, peleteado (con la autorización de Novozymes, Copenhague, Dinamarica) Imágenes de izquierda a derecha: Control, Control + 8mg/g pancreatina, Control + amilasa exógena, Control + 8mg/g pancreatina + amilasa exógena. Las flechas rojas indican formaciones porosas en los gránulos de almidón nativo.
Figura 7: Microscopía electrónica de barrido de una dieta de etapa iniciación de pollos de engorde basada en maíz, peleteado (con la autorización de Novozymes, Copenhague, Dinamarica) Imágenes de izquierda a derecha: Control, Control + 8mg/g pancreatina, Control + amilasa exógena, Control + 8mg/g pancreatina + amilasa exógena. Las flechas rojas indican formaciones porosas en los gránulos de almidón nativo.
Além de aumentar os sistemas endógenos inadequados, há diversos outros benefícios potenciais pela adição de amilase exógena às dietas de aves e suínos. Primeiro, a produção pancreática é responsiva à necessidade e a relativa abundância de diversos enzimas é ajustada para cima ou baixo, dependendo da composição e digestibilidade dos macronutrientes que estão chegando. Hulan & Bird (1972), por exemplo, utilizaram de forma alternada (a cada 4 dias) dietas isonitrogenadas, com concentrações variáveis de gordura e amido, para alimentar pintinhos, e observaram que a atividade amilase e lipase no suco pancreático era ajustada de forma proporcional à ingestão dos respectivos nutrientes. Este fenômeno parece que pode ser aplicado também às enzimas fornecidas de forma exógena. Jiang et al. (2008) observaram uma redução significativa tanto na expressão do mRNA da amilase pancreática como da atividade da amilase endógena em frangos de corte jovens quando a dieta era suplementada com uma amilase exógena. Quando a amilase era adicionada à dieta de frangos de corte, Gracia et al. (2003) observaram aumentos significativos na AMEn, digestibilidade do amido, métrica de desempenho e uma redução na massa pancreática. Estes efeitos são interessantes e oferecem potencial para reduzir os custos de energia e aminoácidos para manutenção através da suplementação das dietas com enzimas exógenas. Estes benefícios na redução das necessidades de aminoácidos para a síntese de amilase podem ser particularmente grandes para glicina e serina, uma vez que a amilase pancreática tem aproximadamente 16-17% Gli+Ser. Ainda que a cooperatividade entre a amilase exógena e endógena não esteja totalmente clara, Pedersen et al. (2015) revelaram algumas evidências relacionadas a isso (Fig. 7).
A microscopia eletrônica de varredura detalhada nos quadros acima (Fig. 7) sugere uma cooperação entre a amilase exógena e a pancreatina para a formação de poros nos grânulos de amido, pelo menos sob condições de teste in vitro (pH 5, 40ºC, 4 h em tampão). Neste trabalho foi observada uma extensa formação de poros e colapso da estrutura dos grânulos de amido, mas somente quando a pancreatina era usada juntamente com a amilase bacteriana exógena e vice versa. O assim chamado ‘amido resistente’ pode ser refratário devido a restrições associadas com acesso físico (RS1), forma cristalina /solubilidade (RS2) ou amido retrógrado após processamento hidrotérmico (RS3) (Tester et al. 2004). É pouco provável que estas restrições na digestão do amido sejam solucionada por mudanças na produção pancreática, podendo haver necessidade de adicionar amilases adicionais (ou mesmo outras enzimas) à ração. Recentemente, Schramm et al. (2016) observaram um aumento significativo (75% vs. 81%) na digestibilidade da ração resistente do amido em uma dieta à base de milho/soja, quando esta dieta foi administrada a pintinhos com ou sem amilase exógena. Além disso, Stefanello et al. (2016) relataram um aumento de 70 kcal/kg na AMEn com a mesma amilase em uma dieta à base de milho/soja dada a pintos de corte. finalmente, a taxa de digestão do amido pode ser aumentada com amilase exógena, resultando em um aumento da digestão no trato GI proximal e uma redução na disponibilidade do amido para o microbioma do intestino grosso. Weurding et al. (2001) mostraram que a rápida digestão do amido pode levar a uma digestibilidade líquida do amido similar à da digestão lenta do amido, mas propuseram que há diferenças na quantidade de amido digerido em diferentes pontos do intestino e que isto pode ter consequências metabólicas que influenciam a eficiência alimentar.
Liu et al. (2014) também observaram que a taxa de digestão do amido (e este índice com a taxa de digestão da proteína) estava correlacionada com a taxa de CA, sugerindo que a digestibilidade ‘ileal’ do amido pode ser menos importante por si só do que a dinâmica de recuperação do amido em relação à proteína. De fato, Liu & Selle (2015) apresentaram uma meta-análise de 33 tratamentos de dieta em diversos estudos de nutrição de frangos de corte e observaram que a taxa de CA ideal pode ser alcançada associando amido de digestão lenta com proteína de digestão rápida. Este mecanismo não está inteiramente claro, mas pode ser que esteja associado ao metabolismo da energia dos enterócitos intestinais. Ainda que a glicose seja uma fonte de energia mais efetiva para os enterócitos, aminoácidos (sobretudo glutamina) são rapidamente catabolizados como uma fonte alternativa de energia, especialmente na ausência de glicose. Por isso, é possível que se uma amilase exógena mudar de forma substancial o local da digestão do amido para regiões proximais do intestino, isto pode ter um efeito deletério sobre a digestão de aminoácidos, uma vez que alguns podem ser catabolizados pelo trato intestinal. Estes fatores precisam ser melhor elucidados uma vez que c podem cooperar para manter um balanço ótimo entre um intestino bem abastecido e a recuperação de macronutrientes para o crescimento.
O amido não é uma molécula simples e homogênea, existindo uma variedade de formas cristalinas com diversas regiões amorfas, perfis de solubilidade e digestibilidade para não ruminantes. A digestão eficiente do amido depende dos esforços cooperativos de várias enzimas exógenas e endógenas, e estes efeitos são influenciados pela composição da ração e o estágio de crescimento do animal. A adição de amilase exógena a dietas de suínos e aves resulta em alterações na função pancreática, que pode trazer vantagem para o animal em termos de custos de manutenção, bem como melhorias substanciais na retenção de amido resistente. Acelerar a taxa de digestão do amido, no entanto, pode não trazer efeitos benéficos sempre para o desempenho do animal, particularmente em casos onde as taxas de digestão de proteína são lentas. O potencial para efeitos cooperativos entre amilase e protease exógenas é considerável para assegurar sincronicidade entre digestão de proteína e amido para um ótimo desempenho.
28 August 2018
We detected that you are visitng this page from United States. Therefore we are redirecting you to the localized version.



