


-
English
- Global
- North America
- Latin America
Ao eclodir, o pintinho não está adequadamente preparado para digerir eficientemente a dieta. Isso, associado aos níveis e à composição inconstantes dos polissacarídeos não amiláceos (PNA) durante o período de produção, torna necessário ajustar a suplementação enzimática.
Os nutricionistas que trabalham com aves enfrentam o desafio de fornecer nutrição adequada ao sistema intestinal imaturo do pintinho durante a fase inicial. Nos dias que sucedem a eclosão, a ave jovem apresenta limitações fisiológicas relativas à quantidade de energia, aos aminoácidos e a outros atributos nutricionais que podem obter de dietas de alta qualidade (Batal e Parsons, 2002).
Os PNA complicam esse processo por restringir o acesso aos nutrientes e interferir na digestibilidade. Esse problema é maior especialmente nas aves mais jovens, até certo ponto, pelo fato de os efeitos impostos pelos PNA serem mais pronunciados nos pintinhos mais novos.
Mesmo assim, a digestibilidade abaixo do ideal pode advir de fatores que vão além da imaturidade intestinal ou da presença dos PNA. Ingredientes de baixa qualidade, enfermidades, sobrecarga de amido e outros fatores podem aumentar o custo da dieta.
É claro que o crescimento inicial é vital para o desempenho subsequente e para a produção de carne (Ross, 2009). As células satélites — incrivelmente dependentes da nutrição inicial — definem como ocorrerá o desenvolvimento muscular nas aves adultas (Halevy et al., 2003).
A formação celular é rápida e transitória nos primeiros dias de vida, e deficiências nutricionais nessa faixa etária podem desvirtuar o desempenho pelo resto da vida e limitar a produção de carne (Noy e Sklan, 1999; Halevy et al., 2001).
Carboidrases, proteases e fitases podem, potencialmente, resolver alguns dos problemas relativos à digestibilidade e melhorar a utilização do substrato (Adeola e Cowieson, 2011). Vários nutricionistas usam diversas enzimas; porém, muitas vezes não estão seguros sobre qual seria a combinação adequada e de que forma calcular o valor dessas ao usar software de formulação que calculam níveis de inclusão com base no custo mais baixo.
Carboidrases são importantes para os PNA por contribuírem para a energia metabolizável (EM) — o principal influenciador do custo da dieta.
A digestibilidade dos PNA solúveis associados a camas molhadas, cloacas pastosas e outros problemas de desempenho em dietas baseadas em trigo e aveia é visivelmente beneficiada pelas enzimas xilanase e glucanase. No entanto, quando comparadas a essas dietas, as que usam milho/farelo soja são muito mais refratárias às carboidrases (Cowieson, 2010; Slominski, 2011). O milho e o trigo contêm teores similares de PNA; mesmo assim as ineficiências nutricionais são vagas pelo fato de o milho apresentar limitações de desempenho menos óbvias.
A efetividade das carboidrases é grandemente afetada pelo acesso aos PNA ou pela proximidade física da enzima ao PNA. Ao sequencialmente fracionar a parede celular do farelo de soja para expor os componentes PNA, Ouhida et al. (2001) e outros relataram que os PNA aumentam significativamente a degradação enzimática de paredes celulares intactas.
Em uma revisão anterior (Feedstuffs, jan. 27), qualquer que seja o grão, os PNA são compostos complexos formados por diferentes estruturas e ligações químicas que se sobrepõem e entrelaçam. A mera densidade da matriz da parede celular pode impedir que a penetração enzimática ocorra na fração mais central; assim sendo, uma estratégia prudente seria promover a degradação sistemática por diversas enzimas. Enzimas puras clonadas com uma atividade dominante podem não ser eficientes na degradação dos PNA do farelo de soja ou de outros cereais, e a exposição dos componentes dos PNA apenas favorece a degradação (Huisman et al., 1999).
A trituração, o condicionamento e a peletização melhoram a exposição dos PNA às enzimas, assim como o faz a ação da moela. A solubilidade dos PNA, a presença de ramificações e a complexidade de diversos tipos de fibras de PNA advindas de diferentes ingredientes tornam a seleção do mix enzimático um fator crucial.
Em ensaios com aves avaliando xilanases, glucanases ou uma combinação de proteases, amilase e xilanase, Slominski (2011) notou que a falta de resposta por parte das aves às dietas com base em milho/soja indica que parece ser necessário envolver um grupo mais diversificado de enzimas de PNA.
Uma pesquisa da Universidade de Manitoba determinou que a complexidade do mix enzimático estava altamente correlacionada ao melhoramento da digestibilidade proteica no íleo e à relação dieta:ganho (Meng et al., 2005). Foram identificados dois fatores simples, porém importantes: (1) é essencial haver um grupo adequado de enzimas, e (2) as combinações enzimáticas devem necessariamente visar um PNA específico.
Portanto, as enzimas que não são adequadamente combinadas com os substratos contendo PNA não produzem benefícios. Como é de se prever, a combinação enzimática que melhor funcionou para soja, canola e ervilhas não produziu os melhores efeitos com trigo pelo fato de os PNA de leguminosas serem significativamente diferentes dos encontrados em cereais (Meng et al., 2005).
Por exemplo, as xilanases serão pouco benéficas com os PNA dos farelos de soja e canola pelo fato de o substrato ser encontrado em pequenas quantidades nas leguminosas.
Outro trabalho concorda com o fato de ingredientes exigirem carboidrases específicas para os PNA (Malathi e Devegowda, 2001). No caso do milho/farelo de soja, combinações de pectinase com hemicelulase ou de hemicelulase com celulase geralmente melhoram a digestibilidade da proteína e matéria orgânica, bem como da EM aparente quando comparado com o controle sem enzimas (Tahir et al., 2006).
A digestibilidade in vitro do milho/farelo de soja aumentou quando sete enzimas foram adicionadas em um mix; no entanto, individualmente apenas a celulase causou o mesmo efeito (Saleh et al., 2004).
Obviamente, as enzimas com atividade desramificadora não devem ser ignoradas (Huisman et al., 1999). A natureza altamente ramificada das arabinoxilanas do milho, bem como das pectinas no farelo de soja, indica a necessidade desse tipo de enzima.
A desramificação melhora a exposição das arabinoxilanas à xilanase e das pectinas à pectinase.
A principal contribuição das proteases da dieta é melhorar a digestibilidade dos aminoácidos e o desempenho (Dozier et al., 2010; Angel et al., 2011; Freitas et al., 2011).
A combinação de proteases e de diversas carboidrases solubilizou mais eficientemente as proteínas e os componentes da parede celular do farelo de soja que qualquer uma das duas individualmente sob concentrações mais altas (Marsman et al., 1997). Poucos trabalhos têm se dedicado ao estudo das proteases associadas a pectinases ou galactosidases em dietas à base de milho/farelo de soja possivelmente pela dificuldade de encontrar pectinases.
Alguns PNA podem quelatar fitatos (Kim et al., 2005), que são fortemente associados a vacúolos proteicos (Bohn et al., 2007). A digestibilidade do fósforo no íleo melhorou de maneira mais efetiva quando foi adicionado um mix de enzimas PNA associadas a fitases, em vez de apenas fitase (Woyengo et al., 2010).
O mix adequado de carboidrase com protease pode permitir uma maior degradação do ácido fítico na presença de fitases.
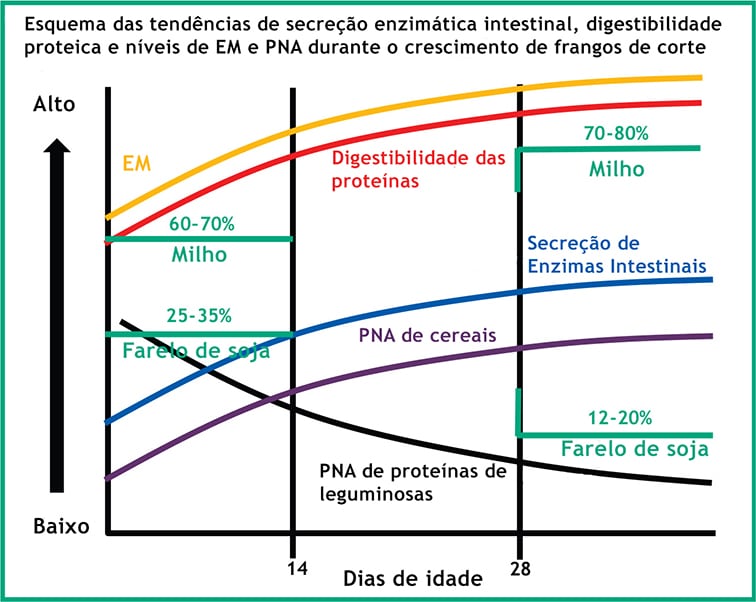
A figura denota a dinâmica que ocorre durante o período típico de recria. Durante esse período, o mix de ingredientes muda naturalmente para atender as exigências nutricionais usando a formulação de custo mais baixo. Essas modificações são acompanhadas por mudanças na composição e nos níveis de PNA, que podem variar de 10 a 15% da dieta.
Ao mesmo tempo, o trato gastrointestinal do neonatal se desenvolve rapidamente. A digestibilidade dos aminoácidos é ruim nos estágios iniciais, comparado aos estados mais avançados, permitindo que uma quantidade significativa da proteína da dieta não seja digerida. Uma vez que o sistema digestivo atinge a maturidade, a digestibilidade dos aminoácidos aumenta, deixando uma menor possibilidade de que proteases exógenas melhorem a digestão.
Desde a ração inicial à de terminação, os níveis de PNA podem variar 30% ou mais. Em uma dieta à base de milho/farelo de soja, por exemplo, os PNA como pectinas e oligossacarídeos do farelo de soja representam uma preocupação maior durante a fase inicial. Na medida em que tem início a fase de recria, a inclusão do farelo de soja na dieta diminui enquanto que os níveis de inclusão de milho e resíduo seco de cervejaria com solúveis (DDGS) aumenta, o que significa que houve um aumento na prevalência de arabinoxilanas.
Conforme ocorre uma alteração no nível do substrato PNA, faz sentido que as principais enzimas PNA também sofrem alteração. O mesmo é verdade para outros substratos, como proteínas, por exemplo, quando proteases podem ter uma maior contribuição no início do período de arraçoamento e o teor de proteína é o mais alto dos estágios de desenvolvimento.
Elaboramos uma série de estudos para suplementar um mix de enzimas adequado à maturidade do trato intestinal enquanto que, ao mesmo tempo, abordamos os níveis e os tipos de substrato durante a vida do frango de corte. As enzimas visam pectinas e oligossacarídeos, bem como PNA de cereais, proteínas, amidos e fitatos.
Experimentos anteriores ajudaram a definir as bases para definir os níveis enzimáticos para enfrentar fatores como idade fisiológica e níveis de substrato na dieta. Essas incluem pectinases, xilanases, amilases, glucanases, enzimas de desramificação, proteases e fitases.
A calorimetria indireta mede o consumo de oxigênio e a produção de dióxido de carbono para quantificar a utilização de nutrientes (McLean e Tobin, 1987). Sistemas de câmaras respiratórias fechadas contendo um animal vivo podem avaliar o gasto energético e quantificar a perda energética resultante de desafios como coccidioses (Teeter, 2010). A calorimetria indireta também permite ter uma boa ideia da eficácia das enzimas para melhorar a EM (Caldas et al., 2014).
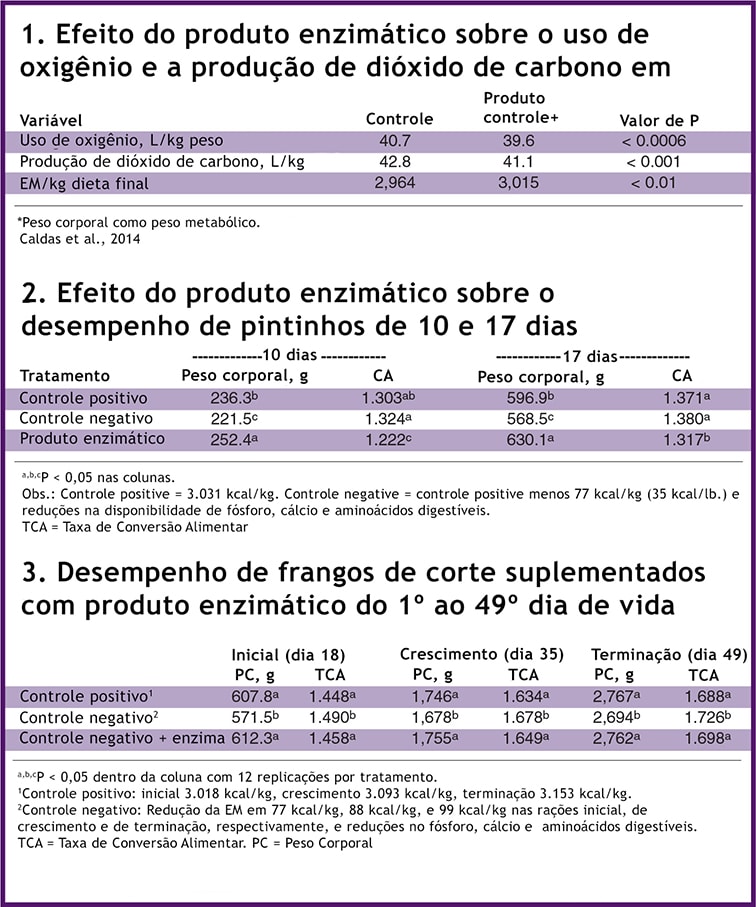
Em um ensaio usando câmaras respiratórias, frangos de corte Cobb receberam uma dieta comercial com e sem enzimas. O produto contendo enzimas baixou (P < 0.0006) o uso de oxigênio metabólico e (P < 0.001) a produção de dióxido de carbono (Tabela 1), indicando que a dieta foi usada com maior eficiência. Em última análise, isso se traduziu em um aumento da ordem de 51 Kcal/kg de EM na dieta final.
Anteriormente, pesquisadores da Universidade Estadual de Oklahoma encontraram uma composição enzimática similar para melhorar (P<0.05) o “valor calórico efetivo” (VCE) da dieta de frango de corte (Teeter et al., 2012) ao usar câmaras respiratórias. A elevação do VCE correspondeu à melhoria (P<0.05) dos pesos corporais e da relação dieta:ganho no mesmo estudo, indicando que houve uma boa concordância entre a pesquisa da câmera de oxigênio e o desempenho das aves nos boxes.
As pesquisas iniciais foram ampliadas para que passassem a incluir baterias de estudos para testar a habilidade desse produto enzimático melhorar o ganho de peso e a conversão alimentar. Em um experimento, pintinhos machos de um dia da linhagem Ross foram alocados aleatoriamente em tratamentos compreendendo dietas básicas à base de milho/farelo de soja contendo 3% de milho DDGS.
O produto enzimático foi adicionado à dieta contendo o controle negativo, que havia sido formulada para conter EM, fósforo e proteína mais baixos em comparação ao controle positivo.
O produto enzimático aumentou o peso corporal em 6,7% e 5,5% e melhorou a relação dieta:ganho em 6,2% e 3,9% nos dias 10 e 17, respectivamente, em frango de corte (Tabela 2).
O grupo controle positivo recebeu uma dieta inicial similar às dietas comerciais típicas; mesmo assim, o desempenho do grupo que recebeu o controle negativo mais enzimas foi superior ao do grupo controle. Isso sugere que, nesse ensaio, a composição enzimática eliminou alguns dos componentes nutricionalmente deletérios da dieta inicial normal.
O desempenho no início da vida é um indicativo do desempenho durante toda a vida, e, aqui, o desempenho no período de 17 dias foi melhor comparado a ambos os grupos controle.
Pesquisa nos boxes dos aviários. A fase final de desenvolvimento focou sobre pesquisas nos boxes dos aviários de frango de corte para testar o produto em condições similares às da prática comercial. Em um experimento, pintinhos de um dia Cobb x Cobb 500 foram aleatoriamente distribuídos a uma taxa de 45 aves por box, havendo 12 replicações por tratamento.
As dietas foram consistentes com as formulações comerciais. As dietas de controle negativo apresentavam déficit de EM, aminoácidos, fósforo e cálcio. Durante toda a vida da ave, o déficit nutricional foi recuperado pela mistura enzimática sem que houvesse a perda de desempenho em termos de peso corporal ou taxa de conversão alimentar (Tabela 3).
Cada fase recebeu uma composição enzimática diferente para levar em consideração tanto o desenvolvimento fisiológico como os substratos disponíveis. Isso evita uma abordagem convencional estática na qual uma enzima ou um grupo de enzimas é usado em todas as dietas, não levando em consideração as mudanças intestinais e de substrato.
Ao eclodir, o pintinho não está adequadamente preparado para digerir os alimentos de maneira eficiente. Esse fato, associado aos níveis inconstantes de PNA e à composição variável da dieta durante o período de produção, torna necessário fazer ajustes à suplementação enzimática. A faixa de componentes de PNA nos ingredientes das dietas e o nível de maturidade fisiológica da ave são variáveis essenciais para se determinar as enzimas a serem usadas na dieta. Ainda não se conhecem todas as sinergias enzimáticas, mas não há dúvidas de que há informações suficientes para se elaborar uma composição mais eficiente de enzimas para dietas à base de milho/farelo de soja visando melhorar a eficiência da produção de carne.
As referências estão disponíveis on-line para membros pagantes em www.Feedstuffs.com ou solicitando diretamente pelo e-mail: tlundeen@feedstuffs.com.
21 August 2018
We detected that you are visitng this page from United States. Therefore we are redirecting you to the localized version.



