


-
English
- Global
- North America
- Latin America
A glicose está no centro do metabolismo energético de aves e suínos e é eficientemente convertida em ATP através do Ciclo do Ácido Cítrico para uso em diversos processos que demandam energia. A maior parte da glicose ingerida por suínos e aves está na forma de amido, geralmente de diferentes grãos de cerais. O amido é um polímero da glicose que consiste em ligações α-1,4 e/ou α-1,6. Quando as ligações α-1,4 ocorrem entre os monômeros da glicose, o polímero resultante é denominado amilose, ao passo que, quando ocorrem ligações α-1,6, o polímero resultante recebe o nome de amilopectina. Por causa da orientação dos monômeros de glicose adjacentes no polímero amilose, desenvolve-se uma hélice natural com uma face externa hidrofílica e um centro hidrofóbico. Estas hélices de amilose formam cristais rapidamente, que são moderadamente insolúveis e razoavelmente inacessíveis para a digestão enzimática. Por outro lado, a amilopectina tem uma estrutura mais caótica uma vez que os pontos de ramificação α-1,6 rompem a estrutura helicoidal, criando regiões amorfas que muitas vezes recebem a denominação “cerosas”. Por isso, as regiões do amido ceroso altamente ramificadas dentro do endosperma dos cereais são muitas vezes um ponto de entrada para que a amilase pancreática e salivar iniciem a degradação do amido que chega a oligômeros dextrina. A relação amilose:amilopectina no amido determina se um amido pode ser classificado como ‘ceroso’ ou ‘rico em amilose’ e isto varia entre os tipos de grãos de cereais e dentro do mesmo tipo. O milho ceroso, por exemplo, pode conter até 99% de amilopectina ao passo que o milho rico em amilose pode ter mais do que 70% de amilose. Além disso, há uma heterogeneidade considerável no tamanho dos grânulos de amido em sementes de diversas plantas e na natureza da microestrutura e da macroestrutura que favorece ou não a digestão. Depois da hidrólise inicial pelas amilases endógenas, o processo de digestão do amido é completado pelas enzimas da borda em escova, maltase e iso-maltase, e a glicose resultante é absorvida por meio de sistemas de transporte dependentes de Na, para eventual metabolismo via o Ciclo do Ácido Cítrico. Como a maioria das dietas de aves e suínos contém 400-500 g/kg amido, a conversão bem sucedida deste amido ingerido em glicose tem uma importância crítica para o status energético do animal, e isto pode ser particularmente verdadeiro em animais mais velhos, com um consumo muito maior de amido. Por isso, fatores que retardam, impedem ou interferem a solubilização e digestão do amido têm um efeito profundo sobre o valor energético de uma determinada dieta. Estes fatores incluem a natureza dos cristais de amido, a extensão do ‘dano’ sofrido pelo amido durante o processamento hidrotérmico da ração ou de seus ingredientes, fatores que podem inibir o funcionamento ótimo das amilases endógenas e questões relacionadas à extração da glicose da luz intestinal pelos sistemas de transporte dependentes de Na. O objetivo deste pequeno artigo é discutir rapidamente estes fatores e delinear a utilidade de fontes exógenas de amilase na nutrição de monogástricos, com ênfase tanto sobre os efeitos diretos da energia digestível como os efeitos indiretos sobre as necessidades de energia líquida e aminoácidos.
O amido é uma estrutura extremamente heterogênea, variando amplamente quanto à composição de amilose e amilopectina, solubilidade, cristalinidade e em sua interação com outros nutrientes, macro e micro, presentes na matéria vegetal (Moran, 1982; Tester et al., 2004). Em geral, os cristais nativos de amido existem como grânulos do tipo A (frequentemente encontrados nos cereais), grânulos do tipo B (frequentemente encontrados em tubérculos) ou grânulos do tipo C (uma mistura dos tipos A e B, frequentemente encontrados nas leguminosas) (Tester et al., 2004).
Os grânulos do tipo A têm uma compactação mais leve e contêm menos água do que os grânulos do tipo B (Wu & Sarko, 1978a, b). Tester et al. (2004) apresentam as características dos grânulos de amido nativo em uma variedade cereais, leguminosas e tubérculos (parte mostrada na in Tabela 1 e Fig. 1), mostrando a heterogeneidade dos tamanhos e formatos dos grânulos que existem em ingredientes comuns, usados em rações para suínos e aves.
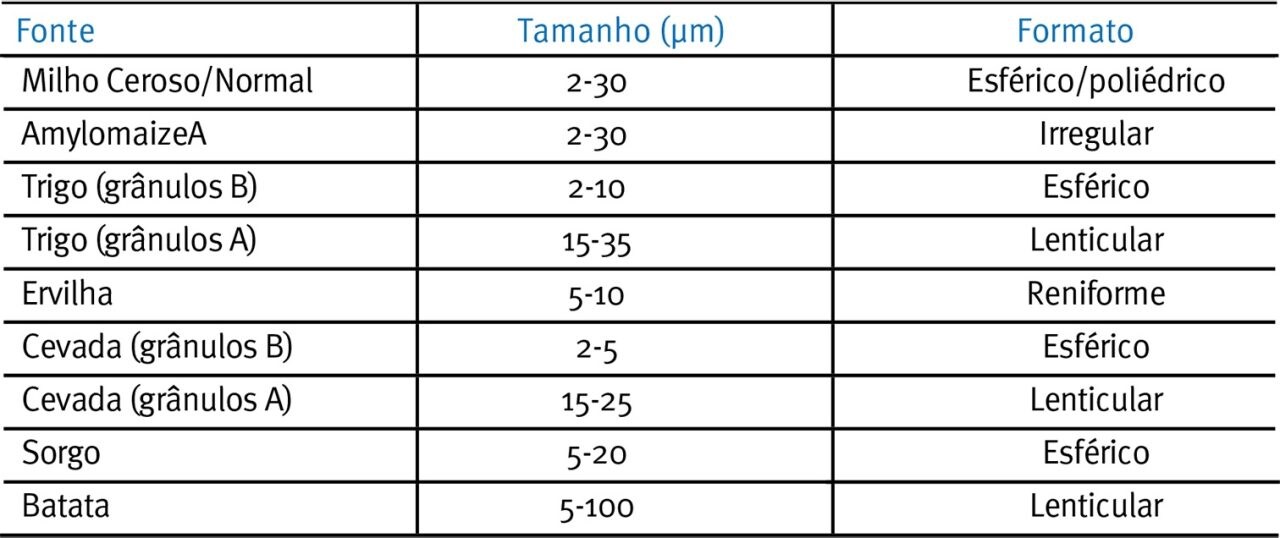 Tabela 1. Características de grânulos de amido nativo de uma variedade de cereais, leguminosas e tubérculos (retirado de Tester et al., 2004).
Tabela 1. Características de grânulos de amido nativo de uma variedade de cereais, leguminosas e tubérculos (retirado de Tester et al., 2004).
 Fig. 1: Grânulos de amido ao microscópio eletrônico de varredura de batata, arroz e trigo (da esquerda para a direita) (Usado com autorização do Prof. E.T. Moran).
Fig. 1: Grânulos de amido ao microscópio eletrônico de varredura de batata, arroz e trigo (da esquerda para a direita) (Usado com autorização do Prof. E.T. Moran).
O amido é definido como ceroso, normal ou amilo quando a razão amilose:amilopectina está em torno de 15%, 16-35% ou >36% respectivamente (Tester et al., 2004; Tabela 2; Fig. 2). Devido aos efeitos de interferência das ligações -1,6 ramificadas da amilopectina sobre a formação de cristais, amidos com uma alta proporção de amilopectina em relação à amilose tendem a ser mais amorfos, solúveis e mais rapidamente digeridos do que os amiloamidos.
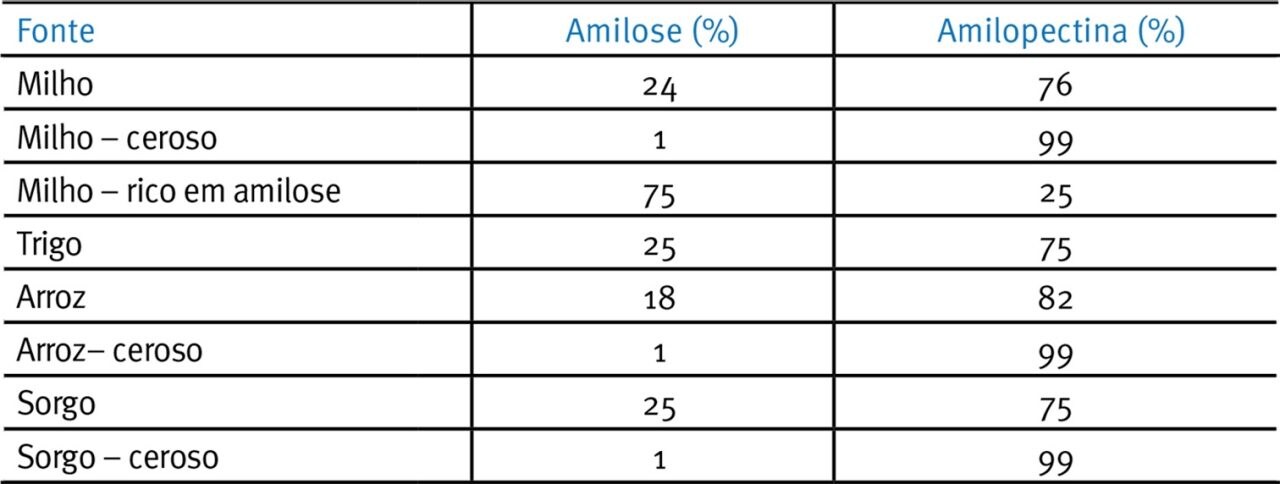 Tabela 2. Composição de amidos e amilose/amilopectina de grãos (Usado com autorização do Prof. E.T. Moran).
Tabela 2. Composição de amidos e amilose/amilopectina de grãos (Usado com autorização do Prof. E.T. Moran).
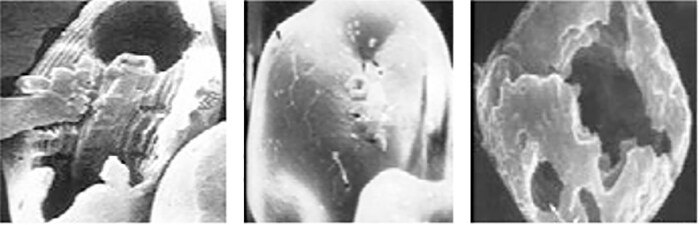 Fig. 2: Grânulos de amido de milho vistos ao microscópio eletrônico: da esquerda para a direita (amilopectina:amilose), normal (75:25), rico em amilose (25:75) ou ceroso (99:1). (Usado com autorização do Prof. E.T. Moran.)
Fig. 2: Grânulos de amido de milho vistos ao microscópio eletrônico: da esquerda para a direita (amilopectina:amilose), normal (75:25), rico em amilose (25:75) ou ceroso (99:1). (Usado com autorização do Prof. E.T. Moran.)
Além disso, nos amidos de cereais apenas os lipídios estão associados aos resíduos de amilose e cerca de 1,5% da massa de grânulos pode ser de lipídios, especialmente nos amidos mais ricos em amilose (Morrison, 1993).
Nas dietas de suínos e aves, globalmente o milho é a principal fonte de amido (Cowieson, 2005) e contribui com a maior parte da energia da dieta. O amido de milho é encontrado no endosperma (cerca de 86%), ainda que cerca de 4-5% de amido seja encontrado no germe e nas camadas de aleurona, respectivamente (Moran, 1982). É importante observar que, no milho, os grânulos de amido são encontrados incorporados em uma matriz proteica que geralmente é hidrofóbica, e pode estar mais ou menos compactados, dependendo da genética e das condições durante a fase de desenvolvimento vegetal.
Quando a matriz amido/proteína do milho está bem compactada e for rica em proteína em relação ao amido, o endosperma é descrito como cristalino ou altamente vítreo, e quando a matriz amido/proteína está pouco compactada e friável, o endosperma é descrito como farináceo (Fig. 3).
 Fig. 3: Milho altamente (A) e pouco (B) vítreo (Gibson et al., 2003), mostrando os grânulos de amido incorporados a uma matriz de proteína prolamina no endosperma.
Fig. 3: Milho altamente (A) e pouco (B) vítreo (Gibson et al., 2003), mostrando os grânulos de amido incorporados a uma matriz de proteína prolamina no endosperma.
Assim, o amido não deve ser considerado como um componente homogêneo independente, mas sim como um composto heterogêneo complexo, consistindo de proporções variáveis de zonas cristalinas e amorfas, lipídio e proteína aderentes e muitas vezes envoltos em uma camada de aleurona e hemicelulose. O sucesso da digestão de amido requer a solubilização das regiões de amilose, predominantemente hidrofóbicas, dissociação do macronutrientes aderidos e a adequação da produção da enzima endógena em relação à capacidade de absorção e o volume de ração que está chegando.
Nos suínos, a digestão do amido é iniciada na boca pela amilase salivar, enquanto que nas aves a digestão do amido somente ocorre quando a amilase pancreática entra em contato com os polímeros de amido ingeridos (Moran, 1985). A α-amilase pancreática hidrolisa a amilose em maltose e maltotriose, e a amilopectina em maltose, maltotriose e dextrinas α-limite, e estes oligossacarídeos são degradados ainda a glicose pela maltase e sucrase-isomaltase na camada não homogênea de água na superfície do microvilo (Moran, 1985). A glicose resultante é transportada através dos sistemas de transporte dependentes de Na para dentro do citoplasma da célula e através da membrana basolateral, para uma eventual entrada no Ciclo do Ácido Cítrico para a geração de ATP. Nas aves, o amido é geralmente digerido quase que por completo (muitas vezes >95%) pelo íleo terminal, mas isto varia e depende da idade, estrutura do amido e solubilidade, processamento hidrotérmico das dietas e outros diversos fatores. Geralmente se considera que as aves desenvolvem uma capacidade maior de digerir amido com o amadurecimento do trato intestinal, e há evidências de uma elevada produção de amilase pancreática nas aves mais velhas, quando comparadas com sua contraparte jovem (Krogdahl & Sell, 1989). De fato, a ave desenvolve a capacidade de digerir amido in ovo (Moran, 1985) e após a eclosão tem uma capacidade superior de digerir amido quando comparada aos mamíferos na fase neonatal, como os leitões que dependem da lactose como fonte predominante de energia. A rápida elevação da produção de amilase pancreática em aves jovens levou à especulação de que esta fase da vida pode ser particularmente responsiva ao aumento dos sistemas de amilase endógena com amilases exógenas microbianas.
No entanto, quando se considera a ingestão de amido (especialmente por unidade de tecido intestinal) são os animais mais velhos e não os mais jovens que podem se beneficiar mais do suprimento de amilase exógena. Croom et al. (1999), por exemplo, observaram que peruzinhos com 2 semanas de idade tinham cerca de quatro vezes mais tecido intestinal por grama de peso vivo do que um peru de 16 semanas de idade (Fig. 4).
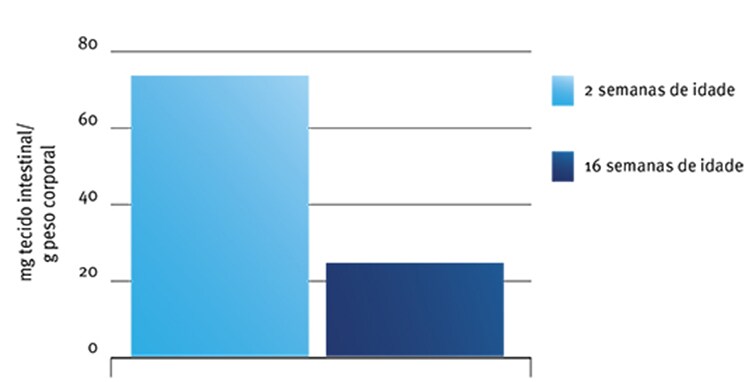 Fig. 4: Efeito da idade sobre o peso intestinal relativo em perus (redesenhado de Croom et al., 1999).
Fig. 4: Efeito da idade sobre o peso intestinal relativo em perus (redesenhado de Croom et al., 1999).
Estes dados, que também são refletidos em frangos de corte, sugerem que o intestino tem uma importância desproporcional no pintinho de corte e o sucesso no desenvolvimento inicial é extremamente importante para assegurar que a ave tenha o suporte adequado em fases posteriores do crescimento. De fato, Lilja (1980; 1983) sugere que o sucesso do desenvolvimento (especialmente para aves com alta capacidade para a taxa de crescimento) em fases posteriores do crescimento está condicionado ao desenvolvimento intestinal adequado no neonato, uma conclusão que é apoiada por Pinchasov et al. (1985).
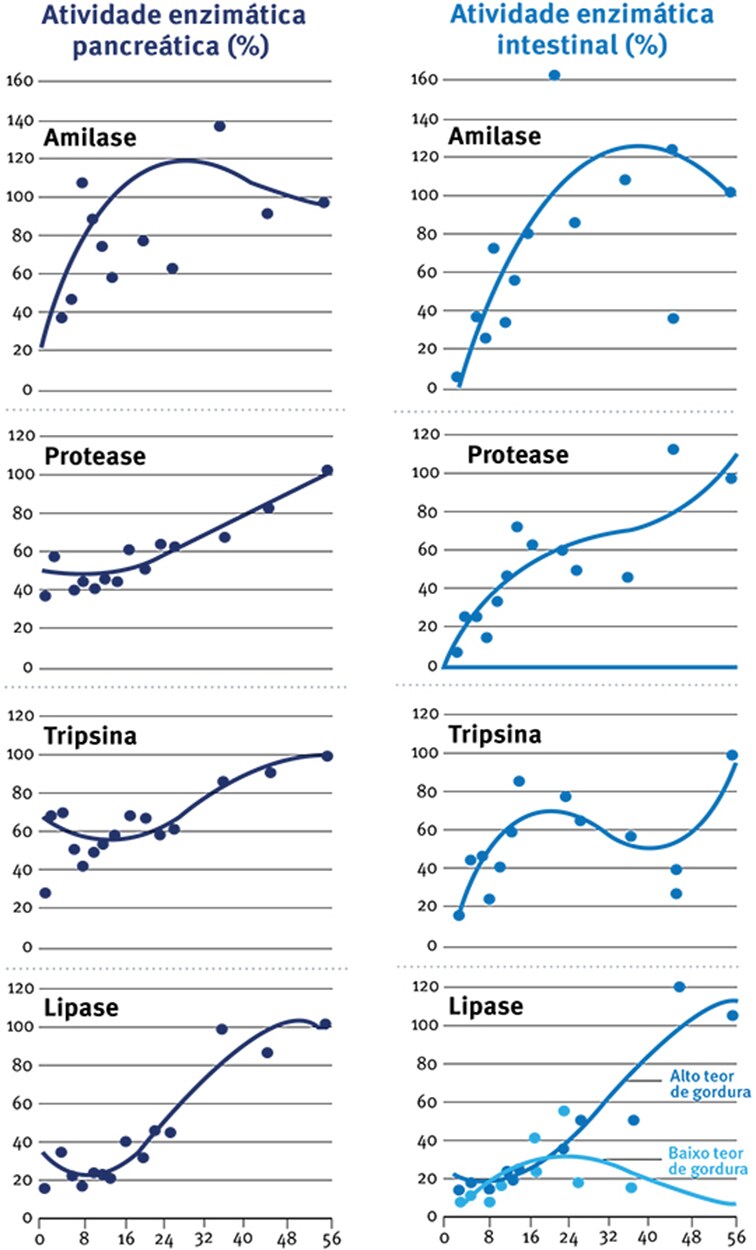 Fig. 5: Desenvolvimento com a idade das enzimas digestivas no tecido pancreático ou intestinal em peruzinhos (expresso como porcentagem dos valores às 8 semanas). (Krogdhal & Sell (1989).
Fig. 5: Desenvolvimento com a idade das enzimas digestivas no tecido pancreático ou intestinal em peruzinhos (expresso como porcentagem dos valores às 8 semanas). (Krogdhal & Sell (1989).
Assim, ainda que a atividade enzimática aumente por si só (Fig. 5) com a idade, em termos de unidades de atividade por grama de tecido pancreático ou intestinal, a quantidade absoluta de enzima digestiva por unidade de peso corporal pode não aumentar. Krogdahl & Sell (1989), por exemplo, mostram que o peso pancreático relativo aumenta de cerca de 0,1% para em torno de 0,5% do dia 1 ao 10, diminuindo depois para cerda de 0,15% no dia 56 (Fig. 6), resultados que estão de acordo com as observações de Croom et al., (1999).
A relevância destas mudanças com o desenvolvimento intestinal em relação ao ganho de peso corporal para o uso de enzimas exógenas como agentes de aumento pode ser contra-intuitivo. Enquanto o pintinho tem claramente uma capacidade limitada de produzir enzimas endógenas, o trato intestinal responde por uma proporção mais substancial de sua massa corporal do que ocorre com o frango de corte na recria/terminação. Assim pode ser mais apropriado que enzimas suplementares, como a amilase, sejam usadas em frangos de corte pesados, e que o intestino e o tecido pancreático passam a ter uma proporção cada vez menor do peso metabólico da ave. Croom et al. (1999) e Krogdahl & Sell (1989) mostram que, ainda que a atividade da enzima intestinal no dia 8 esteja ao redor de 40% da atividade no dia 56 por unidade de tecido intestinal, como porcentagem do peso corporal há aproximadamente 4 vezes mais tecido intestinal em um pintinho do que na ave mais velha. Estes resultados sugerem que a massa intestinal e a atividade das enzimas podem ser mais limitantes nas aves a partir do dia 35 do que nos pintinhos. Ou, pelo menos, que o dito popular segundo o qual os pintinhos precisam somente de enzimas exógenas para melhorar a função digestiva ruim pode não estar totalmente correto.
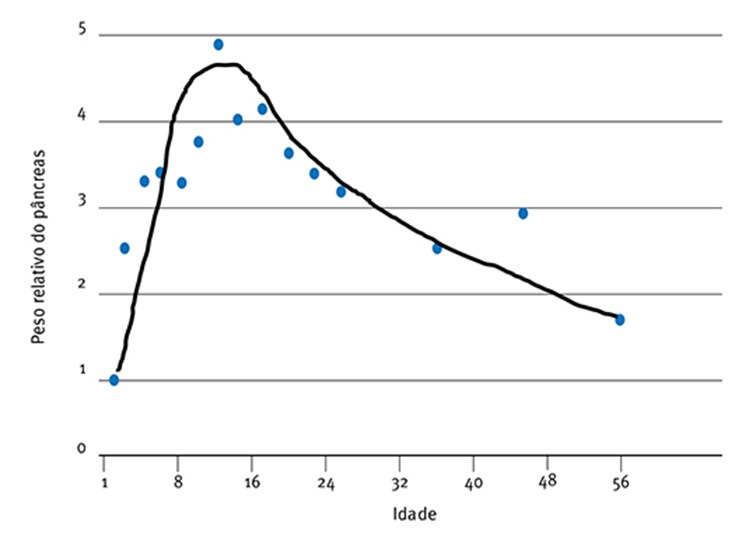 Fig. 6: Desenvolvimento do peso relativo do pâncreas em peruzinhos como porcentagem do peso vivo (Krogdhal & Sell (1989).
Fig. 6: Desenvolvimento do peso relativo do pâncreas em peruzinhos como porcentagem do peso vivo (Krogdhal & Sell (1989).
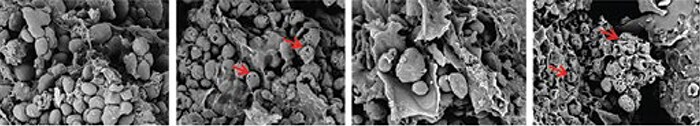 Fig. 7: Imagem obtida com microscopia eletrônica de varredura de uma dieta inicial, peletizada, à base de milho (usada com autorização de Novozymes, Copenhagen, Dinamarca). Os quadros mostram (da esquerda para a direita): Controle, Controle+8mg/g pancreatina, Controle+ amilase exógena, Controle+ 8mg/g pancreatina+ amilase exógena. As setas vermelhas indicam a formação de poros nos grânulos de amido nativo.
Fig. 7: Imagem obtida com microscopia eletrônica de varredura de uma dieta inicial, peletizada, à base de milho (usada com autorização de Novozymes, Copenhagen, Dinamarca). Os quadros mostram (da esquerda para a direita): Controle, Controle+8mg/g pancreatina, Controle+ amilase exógena, Controle+ 8mg/g pancreatina+ amilase exógena. As setas vermelhas indicam a formação de poros nos grânulos de amido nativo.
A microscopia eletrônica de varredura detalhada nos quadros acima (Fig. 7) sugere uma cooperação entre a amilase exógena e a pancreatina para a formação de poros nos grânulos de amido, pelo menos sob condições de teste in vitro (pH 5, 40ºC, 4 h em tampão). Neste trabalho foi observada uma extensa formação de poros e colapso da estrutura dos grânulos de amido, mas somente quando a pancreatina era usada juntamente com a amilase bacteriana exógena e vice versa. O assim chamado ‘amido resistente’ pode ser refratário devido a restrições associadas com acesso físico (RS1), forma cristalina /solubilidade (RS2) ou amido retrógrado após processamento hidrotérmico (RS3) (Tester et al. 2004). É pouco provável que estas restrições na digestão do amido sejam solucionada por mudanças na produção pancreática, podendo haver necessidade de adicionar amilases adicionais (ou mesmo outras enzimas) à ração. Recentemente, Schramm et al. (2016) observaram um aumento significativo (75% vs. 81%) na digestibilidade da ração resistente do amido em uma dieta à base de milho/soja, quando esta dieta foi administrada a pintinhos com ou sem amilase exógena. Além disso, Stefanello et al. (2016) relataram um aumento de 70 kcal/kg na AMEn com a mesma amilase em uma dieta à base de milho/soja dada a pintos de corte. Finalmente, a taxa de digestão do amido pode ser aumentada com amilase exógena, resultando em um aumento da digestão no trato GI proximal e uma redução na disponibilidade do amido para o microbioma do intestino grosso. Weurding et al. (2001) mostraram que a rápida digestão do amido pode levar a uma digestibilidade líquida do amido similar à da digestão lenta do amido, mas propuseram que há diferenças na quantidade de amido digerido em diferentes pontos do intestino e que isto pode ter consequências metabólicas que influenciam a eficiência alimentar. Liu et al. (2014) também observaram que a taxa de digestão do amido (e este índice com a taxa de digestão da proteína) estava correlacionada com a taxa de CA, sugerindo que a digestibilidade ‘ileal’ do amido pode ser menos importante por si só do que a dinâmica de recuperação do amido em relação à proteína. De fato, Liu & Selle (2015) apresentaram uma meta-análise de 33 tratamentos de dieta em diversos estudos de nutrição de frangos de corte e observaram que a taxa de CA ideal pode ser alcançada associando amido de digestão lenta com proteína de digestão rápida. Este mecanismo não está inteiramente claro, mas pode ser que esteja associado ao metabolismo da energia dos enterócitos intestinais. Ainda que a glicose seja uma fonte de energia mais efetiva para os enterócitos, aminoácidos (sobretudo glutamina) são rapidamente catabolizados como uma fonte alternativa de energia, especialmente na ausência de glicose. Por isso, é possível que se uma amilase exógena mudar de forma substancial o local da digestão do amido para regiões proximais do intestino, isto pode ter um efeito deletério sobre a digestão de aminoácidos, uma vez que alguns podem ser catabolizados pelo trato intestinal. Estes fatores precisam ser melhor elucidados uma vez que c podem cooperar para manter um balanço ótimo entre um intestino bem abastecido e a recuperação de macronutrientes para o crescimento.
O amido não é uma molécula simples e homogênea, existindo uma variedade de formas cristalinas com diversas regiões amorfas, perfis de solubilidade e digestibilidade para não ruminantes. A digestão eficiente do amido depende dos esforços cooperativos de várias enzimas exógenas e endógenas, e estes efeitos são influenciados pela composição da ração e o estágio de crescimento do animal. A adição de amilase exógena a dietas de suínos e aves resulta em alterações na função pancreática, que pode trazer vantagem para o animal em termos de custos de manutenção, bem como melhorias substanciais na retenção de amido resistente. Acelerar a taxa de digestão do amido, no entanto, pode não trazer efeitos benéficos sempre para o desempenho do animal, particularmente em casos onde as taxas de digestão de proteína são lentas. O potencial para efeitos cooperativos entre amilase e protease exógenas é considerável para assegurar sincronicidade entre digestão de proteína e amido para um ótimo desempenho. Referências bibliográficas disponíveis mediante solicitação.
09 September 2018
We detected that you are visitng this page from United States. Therefore we are redirecting you to the localized version.



