-
Global/EN
- Global
- North America
- Latin America

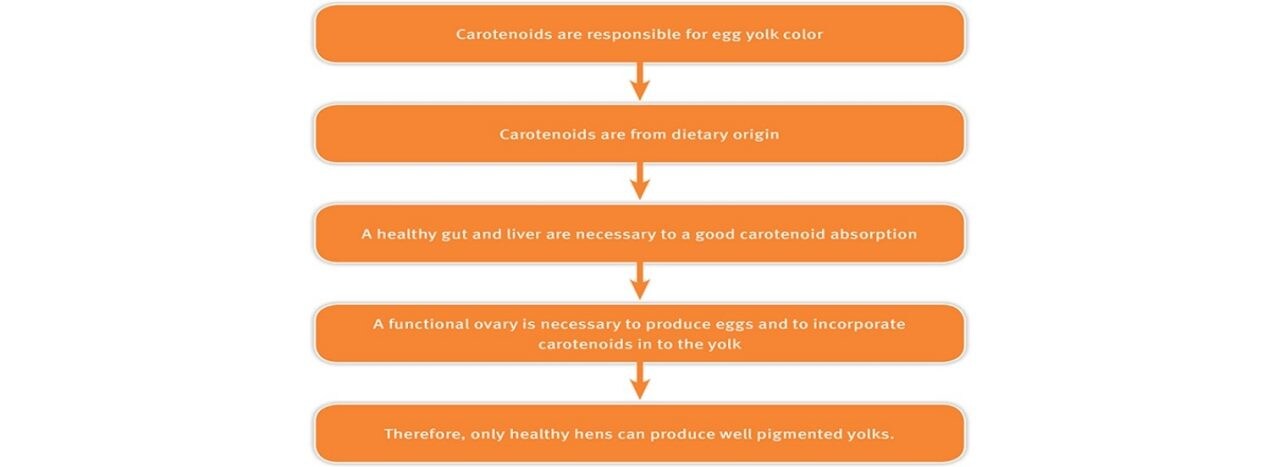
Color is one of the most important factors affecting consumer choices through sensory evaluation of food, including egg yolks. In most countries, golden yolks have been traditionally associated with good health, and despite regional differences on the preferred shade of red and yellow, intense, bright colors are recognized as a sign of a healthy yolk. Moreover, eggs with vivid yolk color can be used to cook attractive pasta, bakery products or sauces. The yolk consists of fats, proteins, vitamins, minerals and carotenoids. Carotenoids are responsible for yolk color, but also required for the development of other physiological functions.
One of the most important of them is a competent immune system, essential for laying hens. There is a trade-off between carotenoids transferred to the egg versus those used to support other functions. Only healthy birds are able to deliver most of feed-derived carotenoids to the egg yolk, therefore a bright yolk color can be considered an indicator of the good health and performance of the flock.
Carotenoids are the most numerous and widespread group of pigments. In 1831, Wackenroder isolated from carrots the crystalline yellow carotenoid then called carotene and in 1837 Berzelius extracted the yellow carotenoids of autumn leaves and named them xanthophylls (Tee, 1992; Karnaukhov, 1990). 100 years later, the number of known naturally occurring carotenoids was about 15 in 1933, 80 in 1948, and rose sharply to about 300 over the next 20 years (Ong and Tee, 1992). Today, the carotenoid family is known to include over 750 compounds (Maoka, 2009) that provide different colors from light yellow to dark red. When complexed with proteins, they can produce green and blue colorations (Ong and Tee, 1992). Historically, carotenoids have been known for their egg yolk pigmenting properties and thought responsible to play specific roles in avian embryonic development (Surai, 2002). Nowadays, a growing body of research shows that when specific carotenoids such as canthaxanthin are added to the diet, they can improve the antioxidant capacity of eggs. (Rosa et al, 2012).
Carotenoids are either carotenes, carotenoids that do not hold oxygen in their molecules, or xanthophylls / oxycarotenoids. Although carotenes are excellent sources of vitamin A , only less than 10% of known carotenoids can be converted into vitamin A. Clearly-defined roles for nonprovitamin carotenoids have still to be established, but evidence is emerging for several important functions (Surai, 2012) including antioxidant activities, cell signaling and transcription factor regulation, promotion of cell differentiation, regulation of cell proliferation.
It seems likely that both sexual display and immune defences can be influenced by carotenoid availability, providing support for the hypothesis that males allocating greater amounts of carotenoids to sexual pigmentation are advertising their superior health (Blount et al., 2003). Therefore, only fit and healthy birds can afford substantial amounts of carotenoids to be directed to the plumage. Any health-related problems (parasites, immunosuppression, oxidative stress, dietary deficiencies) would be associated with increased carotenoid usage for health reasons and fewer carotenoids would therefore be available for display purposes.
Carotenoids absorption from the intestinal tract is associated with the same intraluminal, membrane and intracellular events of dietary lipids. Despite gaps in our understanding of carotenoid absorption in avian species, it seems likely that in poultry, most of the carotenoids are absorbed in the jejunum. Figure 1 (Surai, 2015) shows a general scheme of intestinal absorption of carotenoids.
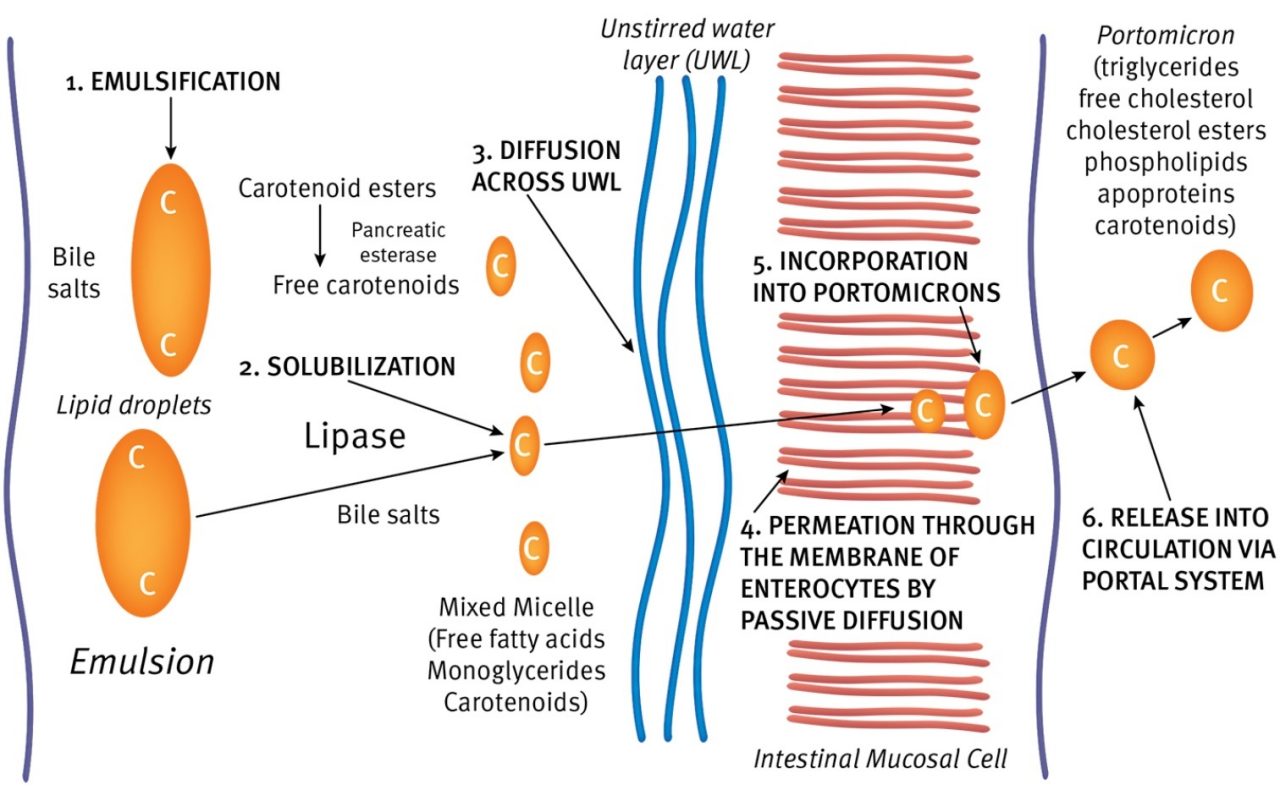 Figure 1. (Surai, 2015) shows a general scheme of intestinal absorption of carotenoids.
Figure 1. (Surai, 2015) shows a general scheme of intestinal absorption of carotenoids.
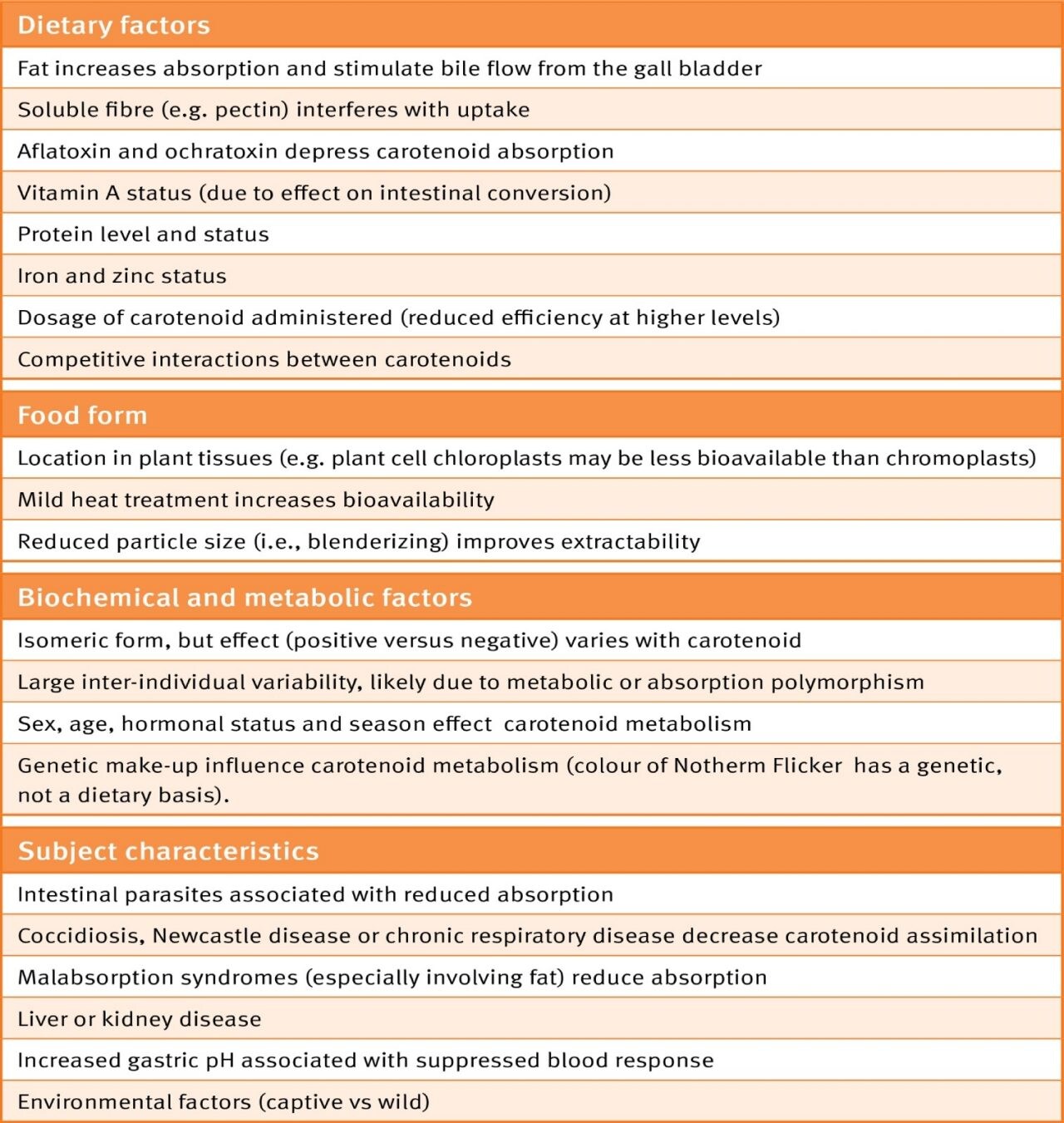 Table 1. Effects of carotenoid absorption and assimilation (Courtesy of Dr. P. Surai, 2015)
Table 1. Effects of carotenoid absorption and assimilation (Courtesy of Dr. P. Surai, 2015)
After the ingestion of feed, carotenoids are released from the matrix by digestive enzymes, including lipase, and further emulsified by bile salts and phospholipids, also involved in the emulsification of dietary triglycerides and other fat-soluble nutrients like Vitamin A, E, K and D3.
The monoglycerides formed during hydrolysis of triglycerides, in the presence of bile salts, spontaneously form very small particles, called mixed micelles. The solubilizing of water-insoluble materials including carotenoids is a critical step in their digestion and absorption. Because of the very small size of mixed micelles, they are dispersed in the aqueous environment of the intestinal lumen and can diffuse into the glycoprotein layer surrounding the microvilli or brush border of mucosal cells, where they come into contact with the cell membranes (Tee, 1992). In general, movement through the unstirred water layer is a rate-limiting factor for lipid absorption (Westergaard and Dietschy, 1976) and on that basis, it appears reasonable to assume that the same is true for carotenoid absorption.
Thus, it is becoming increasingly evident that the micelles formed from dietary lipids serve as a delivery system for carotenoids to reach the absorptive surface of the gut. The micelles facilitate fat absorption by providing a high concentration of lipids in the unstirred water layer adjacent to the mucosal cells.
The feed matrix is thought to be an important determinant for the absorption of carotenoids, as the amount and type of feed determine bile secretion, while bile salts and fat determine micelle formation (van Vliet, 1996). The pH in the intestinal lumen may also affect absorption via an effect on the surface charges of both the micellar particles and the luminal cell membrane, with less diffusion resistance at lower pH (Hollander, 1981). It appears that carotenoids are absorbed by passive diffusion across the brush border membranes of the intestinal mucosal epithelium (Cohn, 1997) and the process is thought to be concentration-dependent (Bieri and Farell, 1976).
It is generally accepted that plasma lipoproteins serve as a transport system not only for lipids but also for carotenoids. The term lipoprotein refers to a set of complex spherical particles formed from lipids and proteins. These particles circulate in the bloodstream, transporting endogenous and exogenous lipids in the plasma (Alvarenga et al., 2011). Indeed, a major function of lipoproteins is to transport lipids through the vascular and extravascular body fluids.
In birds, portomicrons - lipoproteins produced by enterocytes - serve to transport lipids from the gastrointestinal tract to the liver via portal circulation. VLDL (Very Low Density Lipoprotein), LDL (Low Density Lipoprotein) and HDL (High Density Lipoprotein) are the major lipoprotein classes of avian plasma. It is important to understand interactions between lipoproteins and carotenoids to further understand the physiological status of birds. In the liver, some carotenoids are retained by non-parenchymal cells, but the major part is incorporated into newly synthesised VLDL and then released into the bloodstream.
Egg formation is a complex process, involving a substantial increase in lipid and protein metabolism in the liver and other organs. Yolk colour is completely determined by the types and amounts of carotenoids in the hen’s feed and the ability of the hen to absorb and assimilate these pigments.
The hen’s reproductive system is a very complex system that can produce an egg in 24-25 hours. An egg consists of the yolk (30 – 33%), albumen (~ 60%), and shell (9 – 12%). Ovulation usually occurs about 30 minutes after the hen has laid an egg (Lovell et al., 2003). In fact, a female chick is known to be born with a fully-formed ovary containing several thousand tiny ova, or future yolks. When the pullet reaches sexual maturity, the ova begin to develop, one at a time. At any time, an ovary contains ova in various stages of development, ranging from very small, white ova, to almost mature yellow ova, ready to be ovulated. Each yolk is enclosed in its own sac called a follicle. The yolk is kept intact by the surrounding vitelline membrane.
VLDLy (Very-Low-Density-Lipopotein yolk targeted) is the main delivery system for carotenoids into the developing oocyte and it is induced by estrogens (Walzem, et al., 1999). There are some specific unique features of VLDL in laying hens. First of all the diameter of VLDL particles synthesised in the liver of laying hens is about 30 nm, significantly smaller than that in immature hens, and secondly, these so-called yolk-targeted VLDLy are resistant to hydrolysis by LPL (Walzen, 1996). VLDLy small size allows it to pass through the granulosa basal lamina of the ovarian follicle and bind to the LR8 receptor in the oolemma (the membrane that covers the yolk). The receptormediated uptake of intact VLDLy by binding to the follicular apoB receptor for endocytosis (Walzen, 1996) means that carotenoids present in VLDLy will be transported in to the developing follicle.
Therefore, growth of the chicken oocyte is due in large part to the accumulation of yolk, and this has been shown to be due to a receptor-mediated process (Stifani et al. 1990; Schneider 2009). The yolk proteins and lipids are synthesized in the liver through the action of oestrogen, and these proteins are taken into the oocyte by a specific lipoprotein receptor s (LR8; Stifani et al. 1990). Carotenoids are deposited in the yolk to protect the embryo and increase hatchability (Rosa, 2012). Process that could be due to the capacity of carotenoids – such as canthaxanthin - to increase the antioxidant capacity of the egg when fed to the hen (Zhang, 2011).
As consumers in many countries look at golden-yolk eggs as more appetizing and nutritious, it is in the hands of the farmer to offer vivid-coloured eggs to the marketplace. Only healthy birds will be able to produce golden-yolk eggs. As seen with gut infections, any stress or disease-related disturbances to the liver or ovary would decrease carotenoid concentration in egg yolk. The only way to deliver a bright-colored egg is to ensure optimal carotenoid intake and absorption. Healthy birds that receive enough carotenoid will accumulate them in their ovaries and then transfer them to the eggs, for eggs with a consumer-appealing color. A round, shiny and golden yolk always comes from a healthy hen.
References available on request.
28 November 2018

Fernando Cisneros is the Global Layer Solutions Lead. He holds a PhD (University of Illinois) and a MSc (UNAM, Mexico) on animal nutrition, and a BSc on Veterinary Medicine (U. Metropolitana, Mexico).
Fernando was a researcher (INIFAP) and then was hired by Roche Vitamins as technical manager in Mexico and Canada, account and sales manager for DSM Canada, before his current global role. He is passionate about egg quality, bright food and the sustainability of animal farming.
Get in touch with a dsm-firmenich Animal Nutrition & Health specialist or find contacts around the world to suit your needs.
At dsm-firmenich, we love to connect with you.
Follow us on any of the channels below.
We detected that you are visitng this page from United States. Therefore we are redirecting you to the localized version.

