-
Global/EN
- Global
- North America
- Latin America

For producers to raise healthy pigs and to avoid the use of antibiotics, it is critical the piglet has optimal gastrointestinal functionality. However, around weaning there are significant challenges that make this goal difficult to achieve. Optimal gastrointestinal functionality is a situation where the welfare, health and performance of the pig is not constrained by intestinal dysfunction (Celi et al., 2017). To achieve optimal gastrointestinal functionality, there has to be effective digestion and absorption of the feed which requires a normal and stable microbiota, appropriate structure and function of the mucosa and a balanced immune system.
Supporting the piglet at weaning starts with protecting the gut. The gut provides four important functions: it regulates nutrient and fluid uptake, it manages immune tolerance, it provides a defense against infections and it provides a barrier to the external environment. In nature, the piglet is weaned over many weeks in a gradual, prolonged process which permits full development of these functions. However, in commercial production, the weaning process is an abrupt one-day event which brings many challenges to the four essential functions of the gut, culminating in the most commonly reported problem in weaned pigs globally of post-weaning diarrhea stemming from leaky gut syndrome.
Leaky gut syndrome is characterized by hyper-permeability of the intestinal epithelium. A failure in tight-junction functionality results in an increase in paracellular transfer of deleterious compounds (such as bacteria and undigested nutrients) from the intestinal lumen. These deleterious compounds activate the NFkB pro-inflammatory cytokine cascade causing an increased production of reactive oxygen species such as peroxides to fight the microbes but which can also cause collateral damage to cells. Pathogens, such as enterotoxigenic E. coli (ETEC) are then able to adhere to the intestinal cells and produce toxins that induce a loss of water and electrolytes from the cells resulting in diarrhea (Gresse et al., 2017). In addition to diarrhea, the other outcomes of the enteric infection such as reduced feed intake, weight loss, fever, poor feed efficiency and lack of uniformity are easily seen by the producer. What is not easily seen though is the underlying increased production of pro-inflammatory cytokines and acute phase proteins that the enteric infection triggers. The production of these compounds significantly modifies qualitatively and quantitatively the requirement for specific nutrients such as amino acids, vitamins, minerals, antioxidants and lipids.
The severity of leaky gut syndrome is impacted by direct and indirect factors. The direct factors include allergens and antigens that activate the pro-inflammatory cytokine cascade that cause gut inflammation. Allergens can originate from feed and the most prevalent in piglet diets originate from soybean meal. Antigens can also originate from feed and include components such as mycotoxins, but can also originate in the gut lumen from cellular debris (peptidoglycans) originating from cell walls of dead gram-positive bacteria (McCormack et al., 2020).
The indirect factors contributing to the severity of leaky gut syndrome are the facilitators of a gut environment that supports the growth of pathogens both in terms of the substrates they need to grow and the environment in which they live in.
The dual strategy to reduce leaky gut syndrome are outlined in Figure 1.
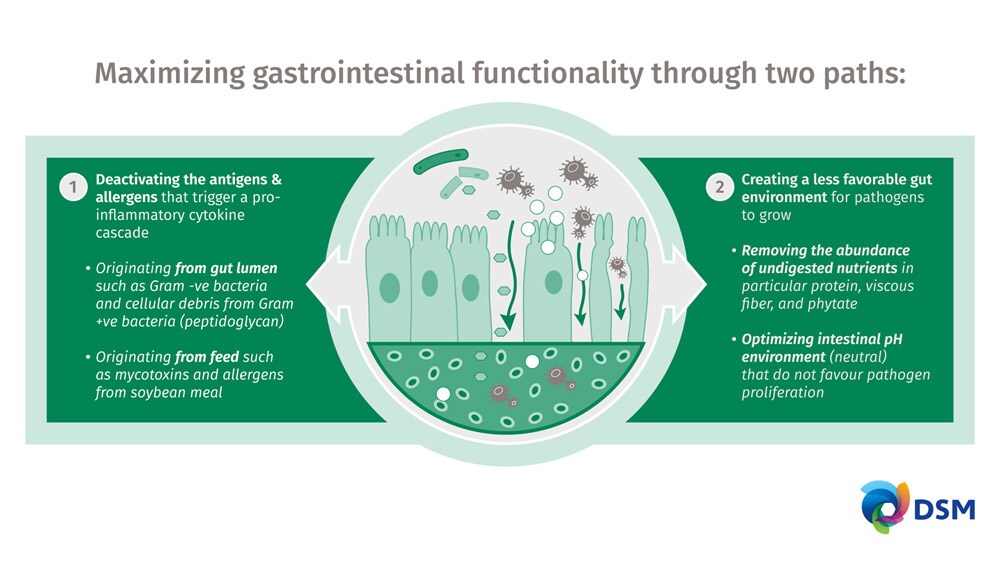
Reducing the need for medications post-weaning relies on protecting the piglet against gut dysfunction that causes diarrhea and supporting optimum gastrointestinal functionality. The achievement of this objective requires a multifaceted approach that is not dependent on a single product strategy but most encompass a comprehensive nutrition approach. In contrast, to tackle this challenge, success lies in the use of tailormade solutions that create a gut environment that is not favorable for pathogen growth in terms of the environment and nutrient supply for the pathogens and deactivating the anti-nutrients arising from the feed and the microbiome.
17 May 2022

Dr. Gannon is an Australian with a Bachelor of Agricultural Science (Honours) and a PhD in pig nutrition from the University of Sydney, Australia. Early in his career, Dr. Gannon worked as a research scientist for university and government organisations in the USA and Australia before moving to the commercial sector in a pig technical and nutrition role with the largest feed miller in Australia. After 10 years, Dr. Gannon started his own nutrition and research consultancy and had a part time appointment as a senior lecturer at the University of Queensland, Australia. In 2011, Dr Gannon joined BIOMIN and prior to the acquisition of BIOMIN by DSM in 2021, Dr. Gannon was the Regional Product Manager for Gut Performance in Asia. At DSM, Dr. Gannon continues his technical support of customers and sales colleagues and research interests as Global Swine Technical Manager.
We detected that you are visitng this page from United States. Therefore we are redirecting you to the localized version.
