-
Global/EN
- Global
- North America
- Latin America

Glucose is central to energy metabolism in poultry and swine and is efficiently converted into ATP via the Citric Acid Cycle for use in various energy-demanding processes. The vast majority of glucose that is ingested by pigs and poultry is in the form of starch, usually from various cereal grains. Starch is a polymer of glucose that consists of α-1,4 and/or α-1,6 bonds. Where α-1,4 bonds occur between glucose monomers the resulting polymer is called amylose whereas when α-1,6 bonds occur the resulting polymer is referred to as amylopectin. Due to the orientation of the adjacent glucose monomers in the amylose polymer a natural helix develops with a hydrophilic external face and a hydrophobic core. These amylose helixes readily form crystals which are moderately insoluble and fairly inaccessible to enzyme digestion. Amylopectin on the other hand has a more chaotic macro-structure as the α-1,6 branching points disrupt the helical structure creating amorphous regions often referred to as ‘waxy’. Therefore, the highly branched waxy starch regions within the endosperm of cereals is often an entry point for pancreatic and salivary amylase to initiate degradation of the incoming starch into dextrin oligomers. The ratio between amylose and amylopectin in starch determines whether a starch may be categorized as ‘waxy’ or ‘high amylose’ and this varies between and within cereal grain types. Waxy corn, for example, may contain as much as 99% amylopectin whereas high amylose corn starch may be more than 70% amylose. Furthermore, there is considerable heterogeneity in the size of the starch granules in seeds from various plants and in the nature of the crystalline micro- and macro-structure that favours, or not, digestion. Following initial hydrolysis by endogenous amylases the process of starch digestion is completed by the brush-border enzymes maltase and iso-maltase and the resulting glucose is absorbed via Na-dependent transport systems for eventual metabolism via the Citric Acid Cycle. As most poultry and swine diets contain 400-500 g/kg starch, successfully converting this ingested starch into glucose is critically important to the animal’s energetic status and this may be particularly true in older animals with a much greater starch intake. Therefore, factors which delay, impede or interfere with the solubilization and digestion of starch have a profound effect on the energy value of a given diet. Such factors include the nature of the starch crystals, the extent of starch ‘damage’ during hydrothermal processing of feed or feed ingredients, factors that may inhibit the optimal functioning of endogenous amylases and issues around extraction of glucose from the lumen via Na-dependent transport systems. It is the purpose of this short article to briefly discuss such factors and to outline the usefulness of exogenous sources of amylase in monogastric nutrition with emphasis both on direct digestible energy effects as well as indirect effects on net energy and amino acid requirement.
Starch is an extremely heterogeneous structure, varying extensively in amylose and amylopectin composition, solubility, crystallinity and in its interaction with other macro- and micro-nutrients inthe plant material (Moran, 1982; Tester et al., 2004). Native starch crystals exist generally as either type A granules (most often found in cereals), type B granules (mostly found in tubers) or type C granules (a mixture of A and B types, often found in legumes) (Tester et al., 2004).
Type A granules are more tightly packed and contain less water than type B granules (Wu & Sarko, 1978a, b). Tester et al. (2004) present the characteristics of native starch granules in a variety of cereals, legumes and tubers (excerpt shown in Table 1 and Fig. 1) which shows the heterogeneity of granular sizes and shapes that exist in common feed ingredients for pigs and poultry.
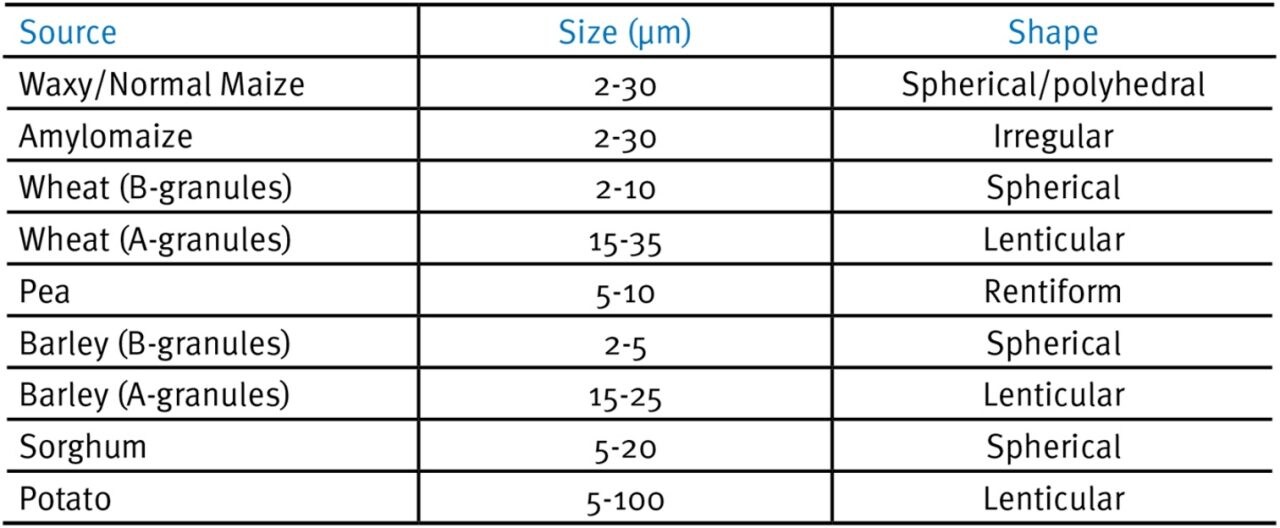 Table 1. Characteristics of native starch granules from a variety of cereals, legumes and tubers (redrawn from Tester et al., 2004).
Table 1. Characteristics of native starch granules from a variety of cereals, legumes and tubers (redrawn from Tester et al., 2004).
 Fig. 1: Starch granules under scanning electron microscopy from (left to right) potato, rice and wheat (Used with permission from Prof. E.T. Moran).
Fig. 1: Starch granules under scanning electron microscopy from (left to right) potato, rice and wheat (Used with permission from Prof. E.T. Moran).
Starch is defined as either waxy, normal or amylo when the ratio of amylose to amylopectin is around 15%, 16-35% or >36% respectively (Tester et al., 2004; Table 2; Fig. 2). Due to the interfering effects of the branched amylopectin α-1,6 bonds on crystal formation, starches with a high proportion of amylopectin relative to amylose tend to be more amorphous, soluble and more readily digested than amylo-starches.
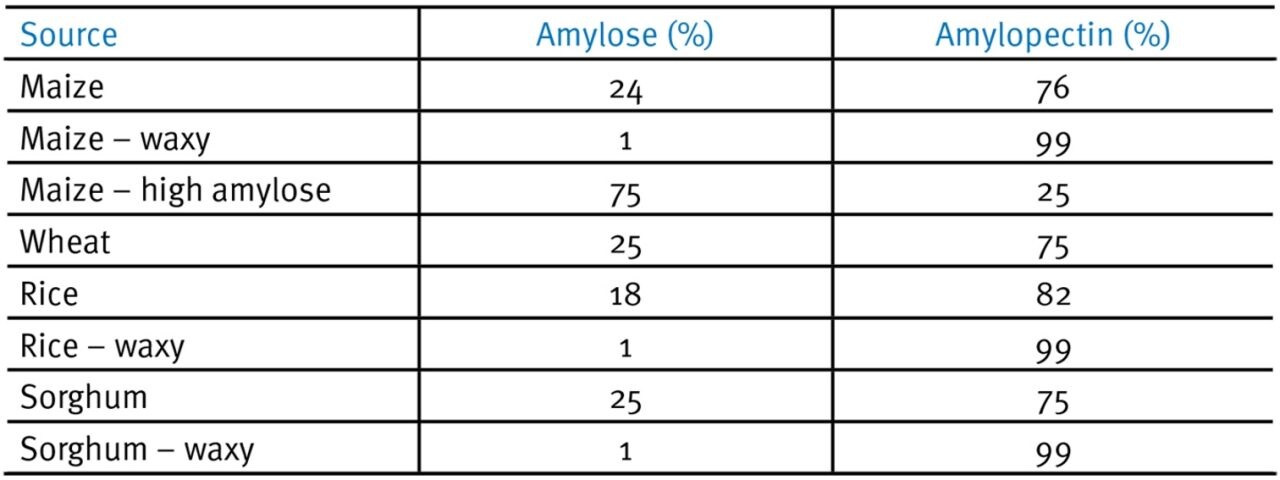 Table 2. Grain starches and amylose/amylopectin composition (used with permission from Prof. E.T. Moran).
Table 2. Grain starches and amylose/amylopectin composition (used with permission from Prof. E.T. Moran).
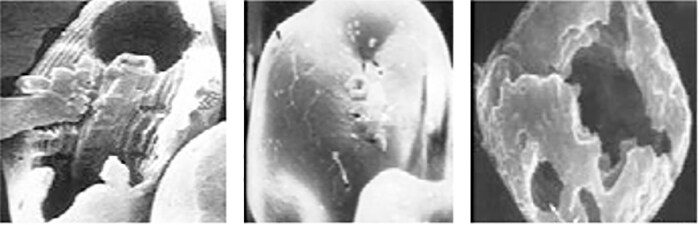 Fig. 2: Corn starch granules under electron microscopy: from left to right (amylopectin:amylose), normal (75:25), high amylose (25:75) or waxy (99:1). (Used with permission from Prof. E.T. Moran.)
Fig. 2: Corn starch granules under electron microscopy: from left to right (amylopectin:amylose), normal (75:25), high amylose (25:75) or waxy (99:1). (Used with permission from Prof. E.T. Moran.)
Additionally, in cereal starches only lipid is associated with amylose residues and around 1.5% of the granule mass may be lipid, especially in higher amylose starches (Morrison, 1993).
Maize is the dominant starch source in the diets of pigs and poultry on a global basis (Cowieson, 2005) and contributes the vast majority of dietary energy. Maize starch is found predominantly in the endosperm (around 86%) though around 4-5% starch is found in the germ and aleurone layers respectively (Moran, 1982). Importantly, in maize, starch granules are found embedded in a protein matrix that is typically hydrophobic and can be either tightly- or loosely-packed depending on genetics and growing conditions.
When the starch/protein matrix in maize is tightly packed and high in protein relative to starch the endosperm is described as flinty or highly vitreous and when the protein/starch matrix is loosely packed and friable the endosperm is described as floury (Fig. 3).
 Fig. 3: High (A) and low (B) vitreous corn (Gibson et al., 2003) showing starch granules embedded in a prolamin protein matrix in the endosperm.
Fig. 3: High (A) and low (B) vitreous corn (Gibson et al., 2003) showing starch granules embedded in a prolamin protein matrix in the endosperm.
Starch digestion in swine is initiated in the mouth by salivary amylase while in poultry starch digestion does not occur until pancreatic amylase comes into contact with ingested starch polymers (Moran, 1985). Pancreatic α-amylase hydrolyses amylose into maltose and maltotriose and amylopectin into maltose, maltotriose and α-limit dextrins and these oligosaccharides are further degraded to glucose in the unstirred water layer by maltase and sucrase-isomaltase at the surface of the micro-villus (Moran, 1985). The resulting glucose is transported via Na-dependent transport systems into the cell cytoplasm and across the basolateral membrane for eventual entry into the Citric Acid Cycle for generation of ATP. In poultry, starch is usually fairly completely digested (often >95%) by the terminal ileum but this does vary and depends on age, starch structure and solubility, hydrothermal processing of diets and various other factors. Fig. 2: Corn starch granules under electron microscopy: from left to right (amylopectin:amylose), normal (75:25), high amylose (25:75) or waxy (99:1). (Used with permission from Prof. E.T. Moran.) 4 Putatively, poultry develop an increased capacity to digest starch as the intestinal tract matures and there is evidence for elevated pancreatic amylase production in older birds compared with their juvenile counterparts (Krogdahl & Sell, 1989). Indeed, fowl develop the capacity to digest starch in ovo (Moran, 1985) and post-hatch have a vastly superior capacity to digest starch compared with neonatal mammals such as piglets who rely on lactose as a dominant energy source. The rapid elevation of pancreatic amylase production in juvenile poultry has led to speculation that this demographic may be particularly responsive to augmentation of endogenous amylase systems with exogenous microbial amylases.
However, when starch intake (especially per unit intestinal tissue) is considered it is actually older animals rather than younger who may benefit most from exogenous amylase supply. For example, Croom et al. (1999) noted that a 2-week old turkey chick had around four times more intestinal tissue per gram of body weight than a 16-week old turkey (Fig. 4).
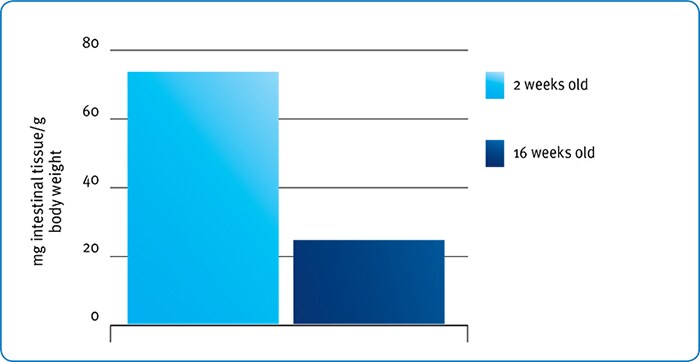 Fig. 4: Effect of age on relative intestinal weight in turkeys (redrawn from Croom et al., 1999).
Fig. 4: Effect of age on relative intestinal weight in turkeys (redrawn from Croom et al., 1999).
These data, which are also reflected in broilers, suggest that the intestine is of disproportionate importance in the young broiler chick and successful early development is extremely important to ensure that the bird is adequately supported in later growth phases. Indeed, Lilja (1980; 1983) suggests that successful development (especially for birds with a high growth rate capacity) in later growth stages is contingent on adequate intestinal development in the neonate, a conclusion that is supported by Pinchasov et al. (1985).
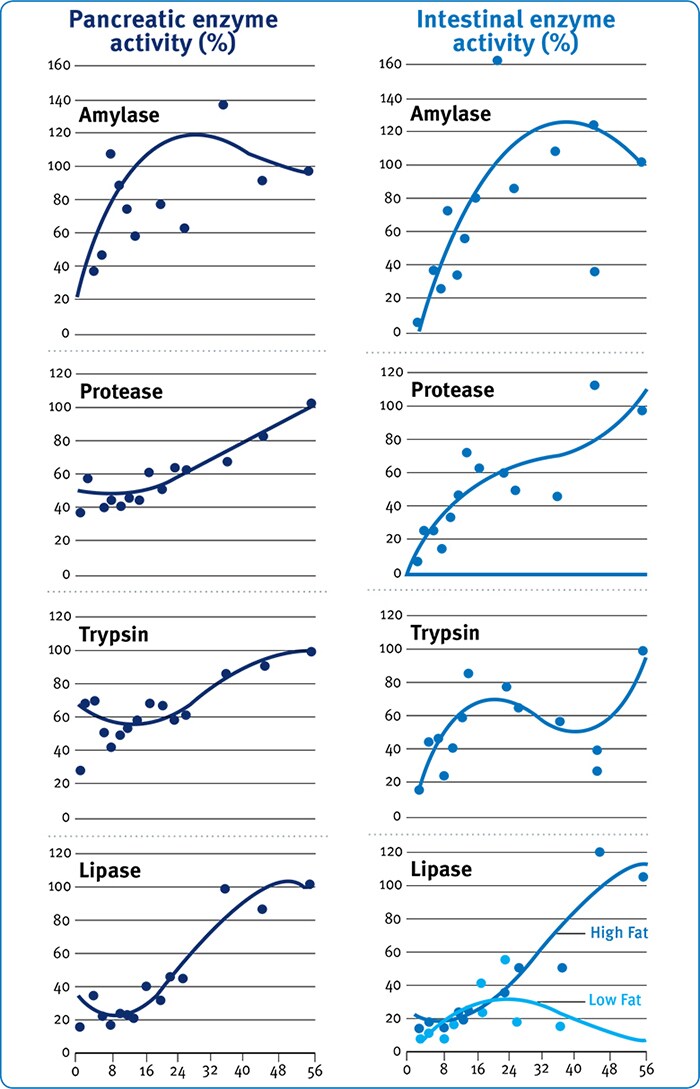 Fig. 5: Development with age (d) of digestive enzymes in pancreatic or intestinal tissue of poults (expressed as a percentage of 8 week values). (Krogdhal & Sell (1989).
Fig. 5: Development with age (d) of digestive enzymes in pancreatic or intestinal tissue of poults (expressed as a percentage of 8 week values). (Krogdhal & Sell (1989).
Thus, though Krogdhal & Sell (1989; Fig. 2) and Uni et al. (1998) show clearly that intestinal function in the neonate is not fully mature until perhaps d14 post-hatch the extent of the inadequacy may not become evident until disproportionate demands are placed on the intestine by the substantial body weight of broilers from d35 to slaughter. In this sense, intestinal and skeletal development have similar consequences where early development is pivotal but consequences of inadequacies may not become evident for several weeks’ post-hatch. Thus, though enzyme activity per se (Fig. 5) increases with age in terms of units of activity per gram of pancreatic or intestinal tissue the absolute amount of digestive enzyme per unit of body weight may not. For example, Krogdahl & Sell (1989) show that relative pancreatic weight increases from around 0.1% to around 0.5% from d1-10 and then decreases to around 0.15% by d56 (Fig. 6), results that are in agreement with observations by Croom et al., (1999).
The relevance of these intestinal developmental changes relative to bodyweight gain for use of exogenous enzymes as augmentative agents may be counter-intuitive in that whilst the young chick has clearly a limited capacity to produce endogenous enzymes the intestinal tract accounts for a more substantial proportion of its body mass than is the case for a grower/finisher broiler. Thus, it may be more appropriate for supplementary enzymes such as amylase to be used in heavy broilers where the intestine and pancreatic tissue become an increasingly diminished proportion of the metabolic weight of the bird. Croom et al., (1999) and Krogdahl & Sell (1989), show that though intestinal enzyme activity on d8 is around 40% of that on d56 per unit of intestinal tissue, as a percentage of body weight there is approximately 4 times more intestinal tissue in a young chick compared with an older bird. These results suggest that intestinal mass and enzyme activity may be more limiting in birds from d35 onwards than in young chicks, or at least that the adage that young chicks uniquely require exogenous enzymes to augment poor digestive function may not be entirely correct.
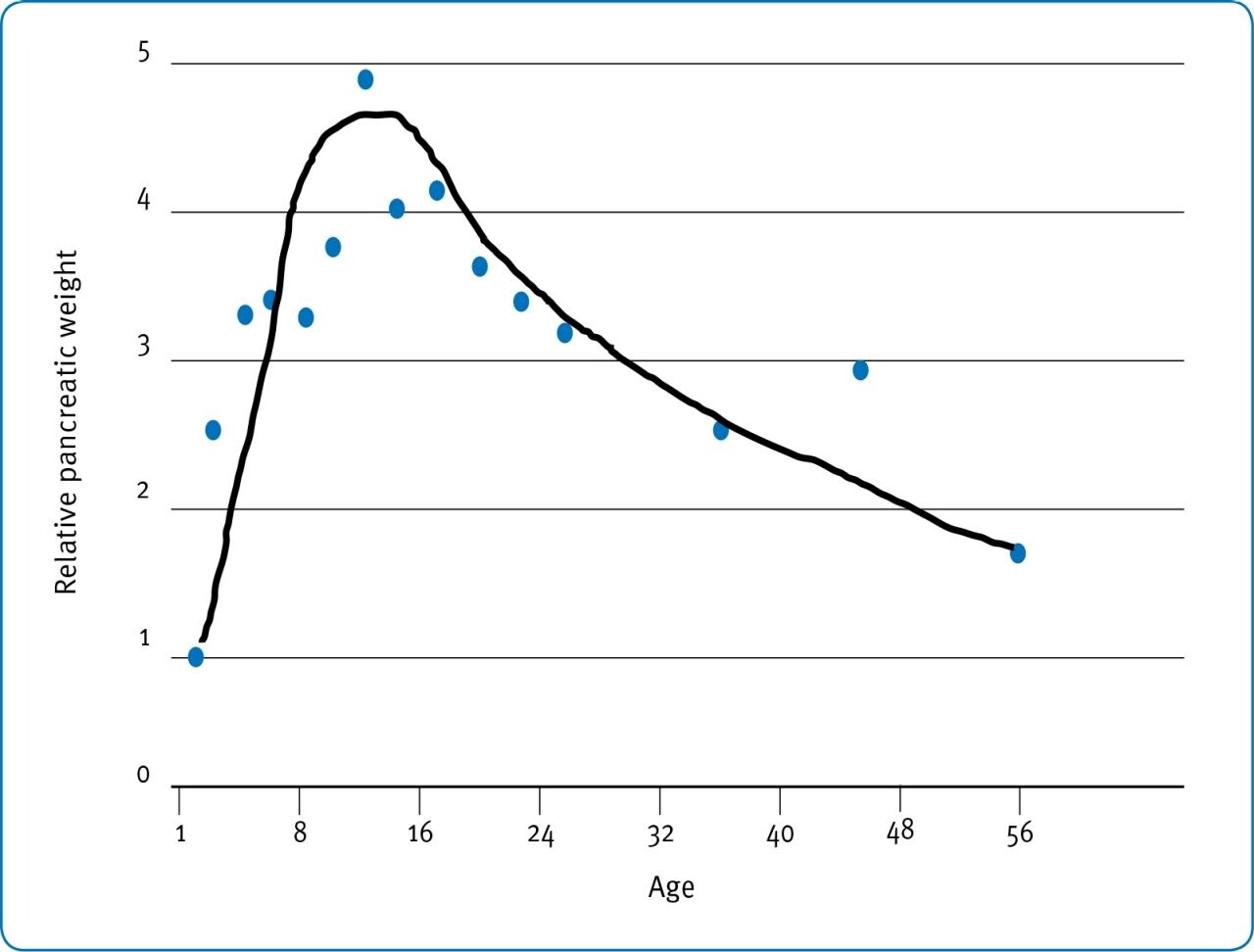 Fig. 6: Development of the relative pancreatic weight in turkey poults as a percentage of live weight. (Krogdhal & Sell (1989).
Fig. 6: Development of the relative pancreatic weight in turkey poults as a percentage of live weight. (Krogdhal & Sell (1989).
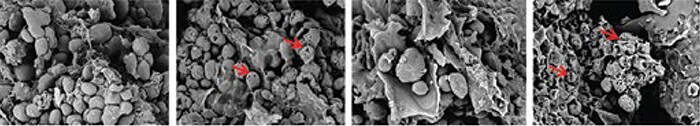 Fig. 7: Scanning electron microscopy of a pelleted corn-based broiler starter diet (used with permission from Novozymes, Copenhagen, Denmark). Frames show (left to right): Control, Control+8mg/g pancreatin, Control+exogenous amylase, Control+ 8mg/g pancreatin+exogenous amylase. Red arrows indicate pore formation in the native starch granules.
Fig. 7: Scanning electron microscopy of a pelleted corn-based broiler starter diet (used with permission from Novozymes, Copenhagen, Denmark). Frames show (left to right): Control, Control+8mg/g pancreatin, Control+exogenous amylase, Control+ 8mg/g pancreatin+exogenous amylase. Red arrows indicate pore formation in the native starch granules.
The scanning electron microscopy detailed in the frames above (Fig. 7) suggest cooperation between exogenous amylase and pancreatin for pore formation in starch granules, at least under the conditions of this in vitro assay (pH 5, 40C, 4hrs in buffer). In this work extensive pore formation and collapse of starch granule structure was observed only when pancreatin was used concurrently with exogenous bacterial amylase and vice versa. Secondly, a portion of the starch in most pig and poultry diets is resistant to digestion. This so-called ‘resistant starch’ may be recalcitrant due to restrictions associated with physical access (RS1), crystalline form/solubility (RS2) or retrograde starch following hydrothermal processing (RS3) (Tester et al. 2004). These restrictions on starch digestion are unlikely to be resolved by changes in pancreatic output and may require additional amylases (or indeed other enzymes) to be added to the feed. Recently, Schramm et al. (2016) noted a significant increase (75% vs. 81%) in the digestibility of the resistant starch fraction in a corn/soy-based diet when this diet was fed to chicks with or without exogenous amylase. Furthermore, Stefanello et al. (2016) observed a 70 kcal/kg increase in AMEn with the same amylase in a corn/soy diet fed to broiler chicks. Finally, the rate of starch digestion may be increased with exogenous amylase resulting in an increase in digestion in the proximal GI tract and a reduction in the availability of starch for the hind gut microbiome. Weurding et al. (2001) showed that rapid starch digestion may lead to similar net digestibility of starch as slow starch digestion but posited that the amount of starch digested at different intestinal sites differs and that this may have metabolic consequences that influence feed efficiency.
Liu et al. (2014) also noted that the rate of starch digestion (and the index of this with the rate of protein digestion) was correlated to FCR, suggesting that the net ‘ileal’ digestibility of starch may be less important per se than the dynamics of starch recovery relative to protein. In fact, Liu & Selle (2015) presented a meta-analysis of 33 dietary treatments over several broiler feeding studies and noted that optimal FCR may be achieved by coupling slowly digestible starch with rapidly digestible protein. The mechanism here is not entirely clear but may be associated with energy metabolism of the enterocytes in the intestine. While glucose is a more effective energy source for the enterocytes in the intestinal tract amino acids (notably glutamine) are readily catabolized as an alternative energy source, especially in the absence of glucose. It is therefore conceivable that if an exogenous amylase substantially shifts the site of starch digestion to proximal regions of the intestine, this may have a deleterious effect on amino acid digestion as some may be catabolized by the intestinal tract. These factors require further elucidation as exogenous amylase and protease may be co-operative to maintain an optimal balance between a well provisioned gut and recovery of macro-nutrients for growth.
Starch is not a simple homogeneous molecule, existing in a variety of crystalline forms with various amorphous regions, solubility profiles and digestibility for non-ruminants. Efficient digestion of starch relies on the co-operative efforts of various exogenous and endogenous enzymes and these effects are influenced by the composition of the feed and the growth stage of the animal. The addition of exogenous amylase to pig and poultry diets results in changes to pancreatic function that may present advantages to the animal in maintenance costs as well as substantial improvements in the retention of resistant starch. However, accelerating the rate of starch digestion may not always have beneficial effects on animal performance, particularly in instances when protein digestions rates are slow. The potential for co-operative effects of exogenous amylase and protease are considerable to ensure synchronicity of protein and starch digestion for optimal performance. References available on request.
28 August 2018

Aaron is a Corporate Science Fellow. He has a PhD in Animal Nutrition obtained from Aberdeen University, UK. Aaron has worked in commercial and academic innovation leadership roles since 2001, most recently as a Professor at the University of Sydney in Australia, joining the DSM innovation team in 2013. Aaron has published more than 200 peer-reviewed papers and book chapters and in addition to his role in DSM, is retained by Purdue University as an Adjunct Professor. In 2016 Aaron was awarded the Poultry Nutrition Research Award by the American Feed Industry Association and in 2019 the Life Mentor Award by the Poultry Science Association.
Get in touch with a dsm-firmenich Animal Nutrition & Health specialist or find contacts around the world to suit your needs.
At dsm-firmenich, we love to connect with you.
Follow us on any of the channels below.
We detected that you are visitng this page from United States. Therefore we are redirecting you to the localized version.

