-
Global/EN
- Global
- North America
- Latin America
At hatch, the chick is not adequately prepared to efficiently digest feed. This, plus the changing levels and composition of NSPs over the production period, necessitates an adjustment in enzyme supplementation.
Poultry nutritionists are challenged with providing proper nutrition to the immature intestinal system of the chick during the starter phase. In the days following hatch, the young bird is physiologically limited in the amount of energy, amino acids and other nutritional attributes it can obtain from high-quality feeds (Batal and Parsons, 2002).
Non-starch polysaccharides (NSPs) complicate this process by encumbering nutrients and interfering with digestibility. This is more problematic at an early age, in part because the effects imposed by NSPs are more pronounced in younger chicks.
Still, suboptimal digestibility can be the consequence beyond intestinal immaturity or the presence of NSPs. Poor quality ingredients, disease conditions, starch overloading and other factors can tax feed costs.
Of course, early growth is vital for subsequent performance and meat yield (Ross, 2009). Satellite cells — remarkably dependent on early nutrition — set the stage for muscle development in the mature bird (Halevy et al., 2003).
Cell formation is rapid and transient in the first days of life, and suboptimal nutrition at this age can pervert lifetime performance and limit meat yield (Noy and Sklan, 1999; Halevy et al., 2001).
Carbohydrases, proteases and phytases can potentially resolve some digestibility issues and improve substrate utilization (Adeola and Cowieson, 2011). Many nutritionists use multiple enzymes but often are unsure of the proper combination and how to best account for their value when using least-cost formulation software.
Carbohydrases for NSPs are important because of their contribution to metabolizable energy (ME) — the primary driver for feed costs.
The soluble NSPs associated with wet litter, pasty vents and other performance issues in wheat- and barley-based diets are visibly responsive to xylanases and glucanases. Compared to these grains, however, corn/soybean meal-based diets are far more reticent to carbohydrases (Cowieson, 2010; Slominski, 2011). Corn contains similar amounts of NSPs as wheat, yet nutritional inefficiencies are vague because performance constraints are less conspicuous with corn.
The effectiveness of carbohydrases is greatly affected by NSP accessibility, or the physical proximity of the enzyme to NSP. By sequentially fractionating cell walls in soybean meal to expose NSP components, Ouhida et al. (2001) and others have reported significant increases in enzymatic NSP degradation relative to the intact cell wall.
Reviewed earlier (Feedstuffs, Jan. 27), the NSP in any grain is a complicated composite of different chemical structures and bonds that overlap and intertwine. The sheer density of the cell wall matrix can hinder enzyme penetration to the inner core; thus, systematic degradation by several enzymes is a prudent strategy. Pure cloned enzymes with one major activity may not effectively degrade NSPs in soybean meal or cereals, and the exposure of NSP components simply favors degradation (Huisman et al., 1999).
Grinding, conditioning and pelleting improves NSP exposure to enzymes, as does gizzard action. Solubility of NSPs, the presence of side chains and the complexity of various types of NSP fiber from different ingredients make the selection of the enzyme mix critical.
In poultry trials with xylanase or glucanase or a combination of protease, amylase and xylanase, Slominski (2011) noted that the lack of response of birds on corn/soybean meal diets indicates that a more diversified group of NSP enzymes seems necessary.
University of Manitoba research found that the complexity of the enzyme mix was highly correlated with improved ileal protein digestibility and feed:gain (Meng et al., 2005). Two simple but important factors were identified: (1) an appropriate group of enzymes is essential and (2) enzyme combinations must target specific NSPs.
Hence, enzymes that are not appropriately paired with NSP substrates exert no benefit. Predictably, an enzyme combination that worked best for soybean meal, canola and peas was not the most optimal for wheat because leguminous NSPs differ significantly from cereal NSPs (Meng et al., 2005).
Xylanase, for example, will serve little purpose with NSPs in soybean and canola meals since the substrate for this enzyme is low in legumes.
Other work concurs that ingredients require carbohydrases specific to the NSP (Malathi and Devegowda, 2001). For corn/soybean meal, pectinase combinations with hemicellulase or with hemicellulase plus cellulase generally showed improved digestibility for protein and organic matter, as well as apparent ME, over the non-enzyme control (Tahir et al., 2006).
In vitro digestibility of corn/soybean meal was improved by a mix of seven enzymes, but individually, only cellulase had the same effect (Saleh et al., 2004).
Certainly, enzymes with debranching side activities should not be overlooked (Huisman et al., 1999). The highly branched nature of corn arabinoxylans, as well as pectins in soybean meal, indicates a need for this type of enzyme. Removing the branches improves exposure of the arabinoxylans to xylanase and the pectins to pectinase.
The primary contribution from feed proteases is improved amino acid digestibility and live performance (Dozier et al., 2010; Angel et al., 2011; Freitas et al., 2011).
A combination of protease and several carbohydrases more effectively solubilized protein and cell wall components in soybean meal than either one did alone at higher concentrations (Marsman et al., 1997). Little work has focused on proteases plus pectinases or galactosidases for corn/soybean meal diets, possibly because pectinases are not widely available.
Some NSPs can chelate phytate (Kim et al., 2005), which is highly associated with protein vacuoles (Bohn et al., 2007). Ileal phosphorus digestibility was more effectively improved with a mix of NSP enzymes plus phytase, as opposed to phytase alone (Woyengo et al., 2010). The appropriate mix of carbohydrases along with protease might permit a greater phytic acid degradation in the presence of phytase.
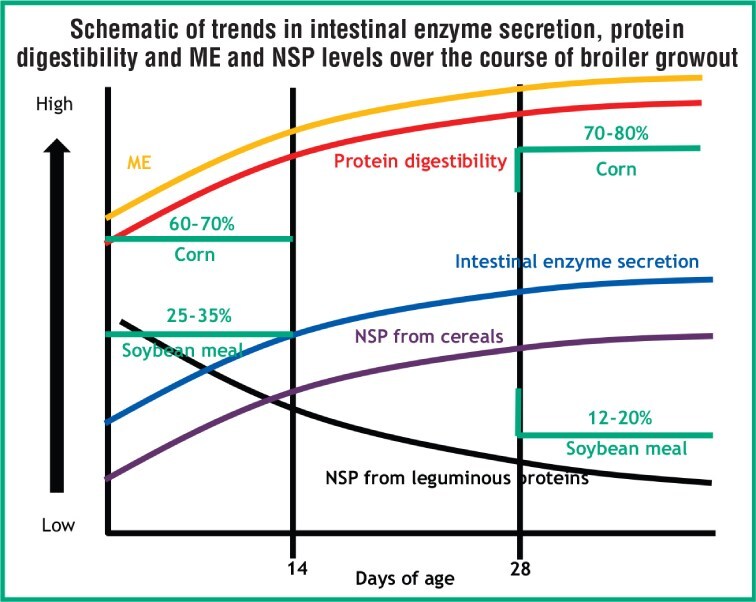
The Figure denotes the dynamics taking place over a typical growout period. During this time, the ingredient mix naturally changes to meet nutrient requirements with a least-cost formulation. These modifications are accompanied by changes in dietary NSP composition and level, which can be 10-15% of the feed.
At the same time, the neonate’s gut is rapidly developing. Amino acid digestibility is poor in early versus older ages, leaving a significant amount of dietary protein undigested. Once the digestive system matures, amino acid digestibility increases, leaving less opportunity for exogenous proteases to improve digestion.
From starter to finisher, NSP levels can change 30% or more. In a corn/soybean meal-based diet, for example, NSPs such as pectins and oligosaccharides from soybean meal are a bigger concern during the starter phase. As the growout ensues, inclusion of soybean meal declines while the inclusion levels of corn and dried distillers grains plus solubles (DDGS) increase, meaning arabinoxylans become more prevalent.
As the NSP substrates change, it stands to reason that primary NSP enzymes should change. The same goes for other substrates such as protein, for example, where a protease could make greater contributions early in the feeding period, when protein content is highest.
We conducted a series of studies to supplement an enzyme mix to address intestinal tract maturity while simultaneously addressing the substrate levels and types over the life span of the growing broiler. The enzymes target pectins and oligosaccharides, as well as cereal NSPs, protein, starch and phytate.
Previous experimental work helped establish the basis behind enzyme levels to address physiological age and feed substrate levels. These include pectinase, xylanase, amylase, glucanase, debranching enzymes, protease and phytase.
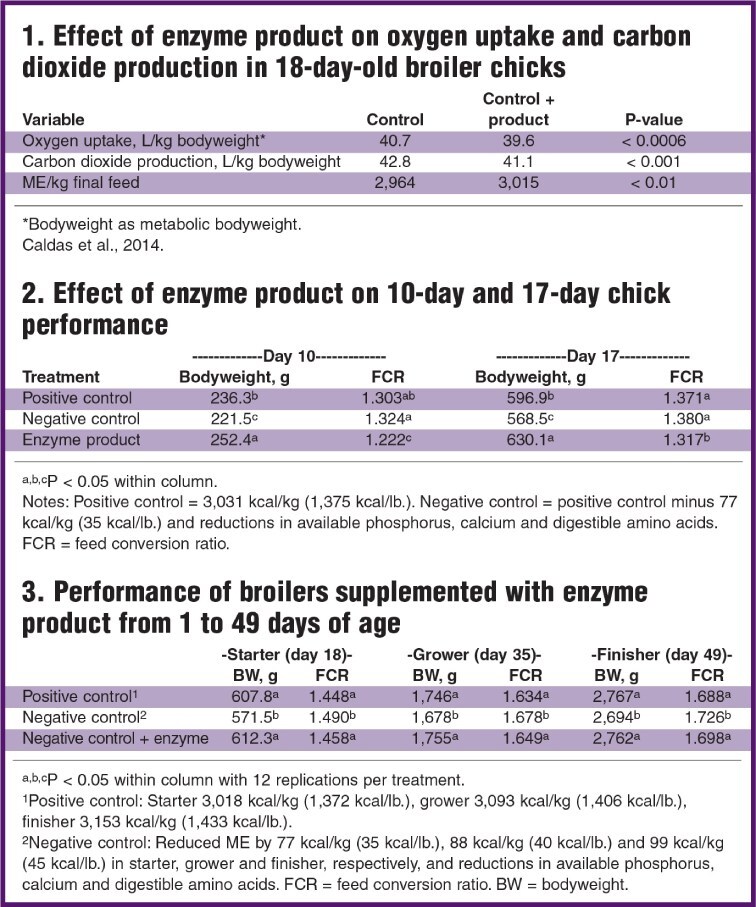
Indirect calorimetry measures oxygen consumption and carbon dioxide production to quantify nutrient utilization (McLean and Tobin, 1987). Closed respiratory chamber systems with a living animal can assess energy expenditures and can quantify energy losses due to challenges such as coccidiosis (Teeter, 2010). Indirect calorimetry also can provide insight on the efficacy of enzymes for improved ME (Caldas et al., 2014).
In a trial using respiration chambers, Cobb broilers were fed a commercial diet with and without the enzyme product. This enzyme product lowered (P < 0.0006) metabolic oxygen uptake and lowered (P < 0.001) carbon dioxide production (Table 1), indicating a more efficient use of the feed. Ultimately, this translated into improved ME by 51 kcal/kg of final feed.
Previously, researchers at Oklahoma State University found a similar enzyme composition to improve (P < 0.05) the “effective caloric value” (ECV) in feed for broilers (Teeter et al., 2012) when using respiration chambers. The elevated ECV corresponded with improved (P < 0.05) bodyweights and feed:gain in the same study, indicating a good agreement between the chamber research and live performance of floor pen birds.
Early research was expanded to include battery studies to test the ability of this enzyme product to improve bodyweight and feed conversion. In one experiment, day old Ross 708 male broiler chicks were randomly allocated across base dietary treatments of corn/soybean meal with 3% corn DDGS.
The enzyme product was added to the negative control diet, which was formulated to be lower in ME, phosphorus and protein compared to the positive control.
The enzyme product increased bodyweight by 6.7% and 5.5% and improved feed:gain by 6.2% and 3.9% on days 10 and 17, respectively, in broiler chicks (Table 2).
The positive control group was fed starter diet similar to typical commercial diets, yet the performance of the negative control plus enzyme group outperformed the control group. This suggests that the enzyme composite eliminated some antinutritional components in the normal starter diet in this trial.
Early life performance is indicative of lifetime performance, and here, 17-day performance was improved over both control groups.
The final phase in development focused on fl oor pen research in broilers to test the product under conditions similar to those in commercial practice. In one experiment, day-old Cobb x Cobb 500 chicks were randomly allocated at the rate of 45 birds per pen, with 12 replications per treatment.
Diets were consistent with commercial formulations. Deficits in ME, amino acids, phosphorus and calcium were present in the negative control diets. Throughout the life of the bird, the nutritional deficit was recouped by the enzyme mixture without performance loss for either bodyweight,or feed conversion ratio (Table 3).
Each phase was provided a different enzyme composite to account for both physiological development and available substrates. This avoids a conventional static approach with one enzyme or group of enzymes across all feeds, which does not account for intestinal and substrate changes.
At hatch, the chick is not adequately prepared to efficiently digest feed. This, along with the changing levels and composition of NSPs over the production period, necessitates an adjustment in enzyme supplementation. Appropriate enzyme selection is crucial to account for the range in NSP components in feed ingredients and the levels of physiological maturity. Not all enzyme synergies are understood, but certainly, sufficient information is available to develop multi-component enzymes for corn/soybean meal diets to improve the efficiency of meat production.
References are available online to subscribers at www.Feedstuffs.com or by request from tlundeen@feedstuffs.com.
05 December 2018
We detected that you are visitng this page from United States. Therefore we are redirecting you to the localized version.

