-
Global/EN
- Global
- North America
- Latin America
While gut health is an increasingly important and popular topic in animal nutrition, a clear scientific definition is still lacking (Kogut and Arsenault, 2016). A definition is needed in order to evaluate the effects of any nutritional intervention on animal health and performance.
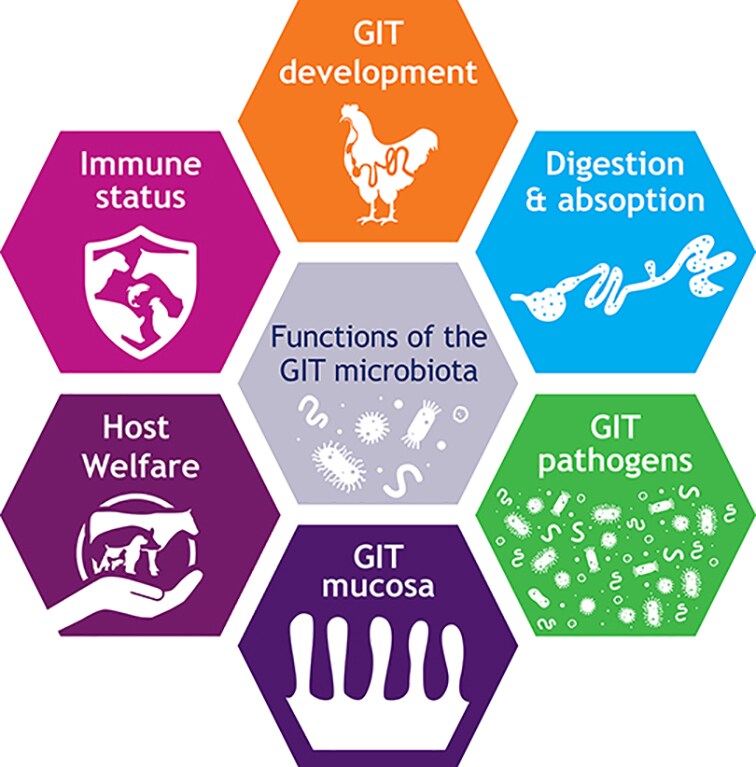 Figure 1. Main components contributing to gastrointestinal functionality.
Figure 1. Main components contributing to gastrointestinal functionality.
We propose the definition of gastrointestinal functionality as “a steady state where the microbiome and the intestinal tract exist in symbiotic equilibrium and where the welfare and performance of the animal is not constrained by intestinal dysfunction” (Celi et al., 2017). This definition combines the principal components of gastrointestinal functionality, namely diet, effective structure and function of the gastrointestinal barrier and normal and stable microbiota, with effective digestion and absorption of feed and effective immune status (Figure 1). All these components play a critical role in gastrointestinal physiology, animal health, welfare and performance. Clarity of understanding of gut health will require the characterization of the interactions between these components.
As the ingested nutrients can play a significant role in the development and functionality of the GIT, diet composition (ingredients, nutrients and additives) can influence the development and function of the digestive system, including the immune system and the microbiota (Conway, 1994).
A large body of evidence has highlighted the potential of using feed supplements, functional foods and ingredients in sustaining animal production performances while maintaining health and welfare (Van Loo, 2007; Pluske, 2013; Starkey, 2014; Hoste et al., 2015). Moreover, the use of functional ingredients seems to support the immune system and to modulate the redox balance and the inflammatory response. The optimization of farm animal diets and nutrition should also consider the various aspects of gastrointestinal functionality that contribute to improvement of animal health and reduction of the use of antibiotics (Allen et al., 2013; Cheng et al., 2014). Therefore, functional properties of feed ingredients and additives related to gastrointestinal health need to play a more central part in diet formulation.
Improved gastrointestinal functionality is often accompanied by improved digestion and absorption, however, one might pose the question whether this is a direct cause or consequence of the observed improvement of GIT functionality (Khadem et al., 2016). Also, one needs to consider that intestinal inflammation negatively impacts GIT function. Indeed, a proliferation in pathogens in the GIT can lead to an increase of defensive responses which is accompanied by a decrease in digestive efficiency and reduced absorption on both macro and micronutrients. Malabsorption is most noticeable for micronutrients, especially iron and zinc (Davin et al., 2012, 2013), while amongst macronutrients, fat is the one that is impacted the most (Koutsos et al., 2003, 2006). Malabsorption can be ascribed to increase in passage rate of digesta along the GIT, hence reducing the time available for nutrients to be digested and absorbed. Malabsorption can also be the consequence of the loss of integrity and function of the GIT barrier, and of a direct response coordinated towards the pathogen by the immune system (Klasing, 2007).
The intestinal microbiota contributes to several physiological functions (Salzman et al., 2007; Lee and Hase, 2014; Marchesi et al., 2016; Round and Mazmanian, 2009) such as protective, structural and metabolic functions. The intestinal microbiota contributes to the regulation of the host homeostasis by contributing to optimal digestion and absorption, regulation of energy metabolism, prevention of mucosal infections and modulation of the immune system.
Consequently, dietary manipulation of the GIT microbiota composition represents an attractive tool to prevent intestinal issues and to promote animal performance. Nutritional interventions should be designed to promote conditions in the GIT that would create and maintain a balance between the host and the GIT microbiota, and to prevent disturbance of the structure and function of the GIT.
What are the factors that influence the development of the GIT microbiota in farm animals? What is the role of the GIT microbiota in the relationships between animal nutrition, GIT physiology, animal health and welfare? While these questions are not easy to answer in a short time, for them to be addressed in their entirety, a multidisciplinary system-based approach must be adopted. It will be imperative to address these questions in order to provide some needed help in the development of alternatives to AGPs. This approach will be crucial to make dietary intervention strategies with new and existing products possible.
The GIT barrier is composed of the mucus layer overlying a monolayer of intestinal epithelial cells and an underlying set of cells composed of mesenchymal cells, dendritic cells, lymphocytes and macrophages constituting the gut associated lymphoid tissue (GALT). Intestinal epithelial cells (IEC) are central in this system as they secrete and regulate the composition of the mucus layer as well as they interact with the underlying cells.
The GIT mucosal epithelium and the associated GALT are the first sites in the host that are exposed to variations in nutrient intake. While, relatively little is known on how nutrients can alter development and function of the GALT, it has been observed that diet can modulate the development of GALT as feeding a casein-based formula to newborn piglets may compromise the development of GALT and systemic immune system.
Nutrients intake and microbiota colonization promotes GALT development, which occurs in parallel with the development of the GIT, resulting in a functional immune system. The GALT promotes gastrointestinal functionality by discerning non-pathogenic and pathogenic antigens and feed-borne threads to the host. This homeostatic orientation is continuously challenged by physiological events and accelerated productive performances which can result in a dysregulation of the immune response in the GIT.
When animals are challenged during harsh environmental conditions (heat stress), poor hygiene, high stocking rates or physiological stages of their productive life (pregnancy and lactation), it is critical to assure that nutrient supply not only meets their requirements but also maximizes GALT function and maintains optimal animal welfare. Functional ingredients can lift animal production performances while maintaining health & welfare. Optimal gastrointestinal functionality is crucial for animal health, their performances and welfare and the environment because it improves feed efficiency, reduces use of antibiotics, and sustains food safety. Therefore, optimization of gastrointestinal functionality will promote sustainable animal production.
While it is clear that feeding a diet that fails to provide adequate amounts of essential nutrients will immune-compromise the animal, there are some crucial questions that need to be addressed to advance our knowledge in dietary immunomodulation. For example, how do specific nutrients modulate the development of the mucosal immune system and what are the critical windows for intervention? What are the relationships between the microbiota and the host’s immune system? It has been proposed that the host and microbiota form a ‘superorganism’ and that “the immune system is not a killer, but a force that shapes homeostasis in the superorganism” (Eberl, 2010). The establishment and the maintenance of homeostasis is dependent on many factors and this brings further support to our multidisciplinary definition of gut health which combines gut physiology, endocrinology, microbiology, immunology and nutrition.
In our opinion, a better insight in the evaluation and in the progression of several GIT conditions, associated with nutritional interventions to optimize gastrointestinal functionality, needs to be made a priority by the scientific community, to translate such knowledge into practical applications, for animal scientists and feed formulators, for ultimately further conferring health and welfare to the animals.
Allen, H.K., Levine, U.Y., Looft, T., Bandrick, M., Casey, T.A., 2013. Treatment, promotion, commotion: antibiotic alternatives in food-producing animals. Trends Microbiol. 21, 114-119.
Celi, P. Cowieson, A.J., Fru-Nji, F., Steinert, R.E., Kluenter, A-M., Verlhac, V. 2017. Gastrointestinal functionality in animal nutrition and health: New opportunities for sustainable animal production. Animal Feed Science and Technology 234: 88-100.
Cheng, G., Hao, H., Xie, S., Wang, X., Dai, M., Huang, L., Yuan, Z., 2014. Antibiotic alternatives: the substitution of antibiotics in animal husbandry? Front. Microbiol. 5, 217.
Conway, P.L., 1994. Function and regulation of the gastrointestinal microbiota of the pig, In: Souffrant, W.B., Hagemeister, H. ((Ed.), Proceedings of the VIth International Symposium on Digestive Physiology in Pigs, EAAP Publication, Dummerstof, pp. 231–240.
Davin, R., Manzanilla, E.G., Klasing, K.C., Perez, J.F., 2012. Evolution of zinc, iron, and copper concentrations along the gastrointestinal tract of piglets weaned with or without in-feed high doses of zinc oxide compared to unweaned littermates. J. Anim. Sc.i 90 Suppl 4, 248-250.
Davin, R., Manzanilla, E.G., Klasing, K.C., Perez, J.F., 2013. Effect of weaning and in-feed high doses of zinc oxide on zinc levels in different body compartments of piglets. J. Anim. Physiol. Anim. Nutr. 97 Suppl 1, 6-12.
Eberl, G., 2010. A new vision of immunity: homeostasis of the superorganism. Mucosal immunol. 3, 450-460.
Hoste, H., Torres-Acosta, J.F., Sandoval-Castro, C.A., Mueller-Harvey, I., Sotiraki, S., Louvandini, H., Thamsborg, S.M., Terrill, T.H., 2015. Tannin containing legumes as a model for nutraceuticals against digestive parasites in livestock. Vet. Parasitol. 212, 5-17.
Khadem, A., Lourenço, M., Delezie, E., Maertens, L., Goderis, A., Mombaerts, R., Höfte, M., Eeckhaut, V., Van Immerseel, F., Janssens, G.P.J., 2016. Does release of encapsulated nutrients have an important role in the efficacy of xylanase in broilers? Poult. Sci. 95, 1066-1076.
Klasing, K.C. 2007. Nutrition and the immune system. Br. Poult. Sci., 48 , 525-537
Kogut, M.H., Arsenault, R.J., 2016. Editorial: Gut Health: The New Paradigm in Food Animal Production. Front. Vet. Sci. 3.
Koutsos, E.A., Calvert, C.C., Klasing, K.C., 2003. The effect of an acute phase response on tissue carotenoid levels of growing chickens (Gallus gallus domesticus). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 135, 635-646.
Koutsos, E.A., Garcia Lopez, J.C., Klasing, K.C., 2006. Carotenoids from in ovo or dietary sources blunt systemic indices of the inflammatory response in growing chicks (Gallus gallus domesticus). J. Nutr. 136, 1027-1031.
Lee, W.-J., Hase, K., 2014. Gut microbiota-generated metabolites in animal health and disease. Nat. Chem. Biol. 10, 416-424.
Marchesi, J.R., Adams, D.H., Fava, F., Hermes, G.D.A., Hirschfield, G.M., Hold, G., Quraishi, M.N., Kinross, J., Smidt, H., Tuohy, K.M., Thomas, L.V., Zoetendal, E.G., Hart, A., 2016. The gut microbiota and host health: a new clinical frontier. Gut. 65, 330-339.
Pluske, J.R., 2013. Feed- and feed additives-related aspects of gut health and development in weanling pigs. J. Anim. Sci. Biotechnol. 4, 1.
Round, J.L., Mazmanian, S.K., 2009. The gut microbiota shapes intestinal immune responses during health and disease. Nat. Rev. Immunol. 9.
Salzman, N.H., Underwood, M.A., Bevins, C.L., 2007. Paneth cells, defensins, and the commensal microbiota: A hypothesis on intimate interplay at the intestinal mucosa. Semin. Immunol. 19, 70-83.
Starkey, J.D., 2014. Triennial Growth Symposium--A role for vitamin D in skeletal muscle development and growth. J. Anim. Sci. 92, 887-892.
Van Loo, J., 2007. How chicory fructans contribute to zootechnical performance and well-being in livestock and companion animals. J. Nutr. 137, 2594s-2597s.
10 October 2018
We detected that you are visitng this page from United States. Therefore we are redirecting you to the localized version.

