-
Global/EN
- Global
- North America
- Latin America
In Brief
Diets contaminated with zearalenone can damage pigs’ immune systems, increase inflammation and reduce nutrient absorption. Knowing the level of ZEN contamination in feed and having a comprehensive mycotoxin risk management strategy is key to running a successful swine operation.

Zearalenone (ZEN) is a mycotoxin produced by Fusarium graminearum, F. culmorum, F. crookwellense, F. equiseti and F. semitectum. This mycotoxin regularly co-occurs with deoxynivalenol (DON), as the same fungi (F. graminearum and F. culmorum) can produce both compounds. ZEN contamination in grains can vary from year to year. According to the dsm-firmenich Mycotoxin Survey ZEN was detected in 40%-71% of swine feed samples over the past nine years (Figure 1).
Once ZEN is absorbed, it is transformed into its major metabolites, α-zearalenol (α-ZEL) and β-zearalenol (β-ZEL). In pigs, the main metabolite is α-zearalenol. The toxicity of this metabolite is much higher, which explains the higher sensitivity of pigs to ZEN. The effects of ZEN depend on the amount ingested, length of exposure, age and health status of the animal.
ZEN is particularly known for the reproductive problems that it can cause. Due to its similarity with estrogens, ZEN binds to estrogen receptors (ERs) and disturbs hormonal balance. However, ZEN can also induce oxidative stress, affect major immune organs and interfere with gut immunity.
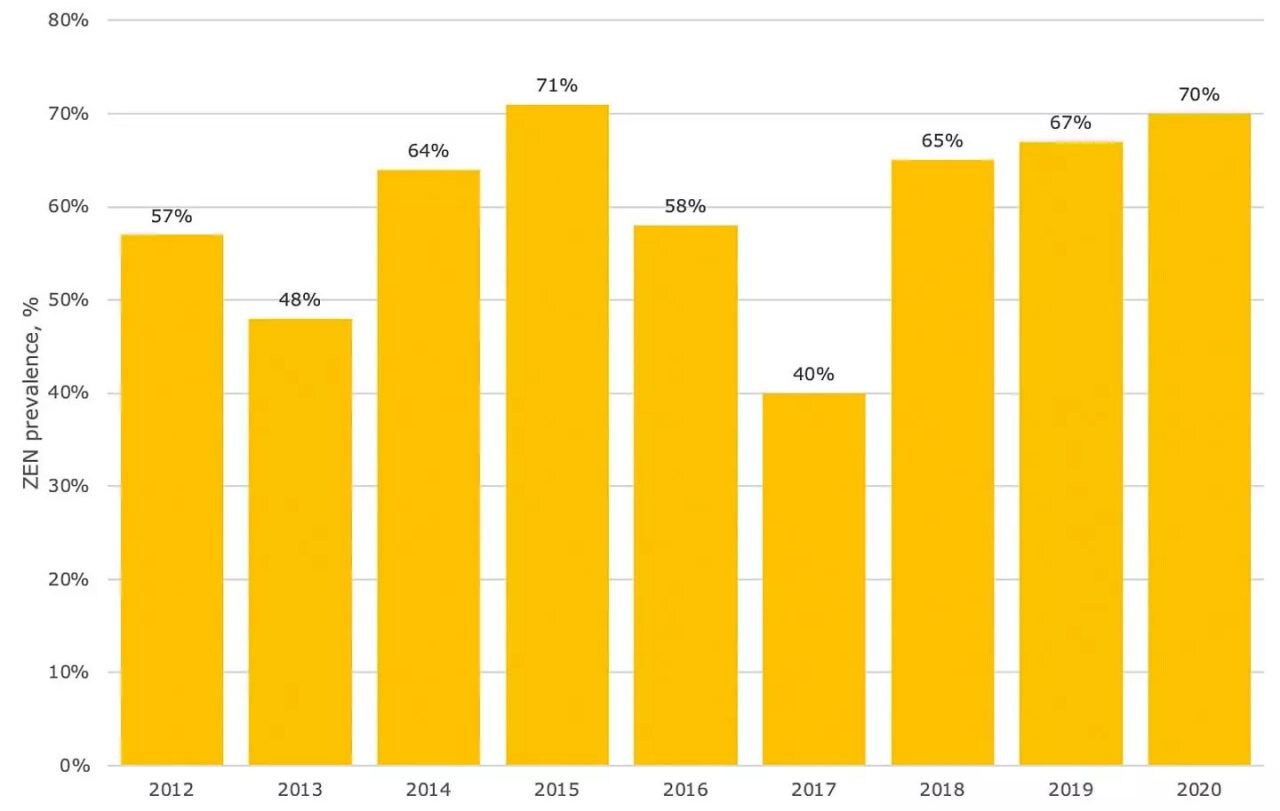
Figure 1. Prevalence of ZEN in final feed worldwide intended for swine. Source: dsm-firmenich Mycotoxin Survey
The effects of ZEN depend on the amount ingested, length of exposure, age and health status of the animal.
Neutrophils are a major part of innate immunity defense. Once inflammation is induced after a challenge, they migrate to the inflammation site leading to phagocytosis by producing reactive oxygen species. An in vitro study (Marin et al. 2010) showed that ZEN and its derivatives have a negative effect on neutrophils’ functionality by decreasing their proliferation and IL-8 and O2- production. In addition, ZEN metabolites have a greater effect compared to ZEN.
In a later in vivo study conducted by the same group (Marin et al., 2013), the effect of ZEN on oxidative stress and inflammation was assessed. Weaning piglets were challenged with ZEN (316 ppb) for 18 days. ZEN acted in a tissue-dependent manner. In the liver, ZEN exhibited inflammatory suppression resulting in hepatic disfunction. In contrast, ZEN induced inflammation in the blood and spleen.
The different impact of ZEN on the spleen and the liver has been demonstrated in another two studies (Pistol et al., 2014; 2015). Piglets intoxicated with ZEN (316ppb) for 18 days showed different immune responses. In the spleen, ZEN triggered an inflammatory response by increasing the synthesis and expression of pro-inflammatory cytokines. In contrast, ZEN induced hepatotoxicity by suppressing the inflammatory response in the liver.
Skiepko et al (2020) evaluated the effect of ZEN, DON and the combination of both ZEN and DON on liver histology and morphological cellular features. In a six-week experimental period, piglets were challenged with ΖΕΝ (40 μg/kg body weight (BW) per day). Hepatocytes’ ultrastructure was affected and iron deposits were elevated. Both changes indicate intoxication and have been related with chronic liver diseases.
Chen et al (2017) assessed the effects of ZEN different levels (1.1 to 3.2 ppm in feed) of immunological parameters in the spleen of young gilts. ZEN ingestion resulted in tissue damage and a decrease in the lymphocyte proliferation rate. The ingestion of ZEN also induced alterations in the production and expression of cytokines.
Wu et al (2020) evaluated piglets that were fed increasing levels of ZEN (ranging from 200 to 1,600 ppb) for 14 days. As expected, ZEN induced hyperestrogenism and alterations in genital hormone levels. Moreover, ZEN reduced antioxidant capacity which is an indicator of the antioxidant response against free radicals produced in the body. In addition, liver enzyme (aspartate aminotransferase) levels increased in blood, indicating liver damage.
ZEN may elicit local effects by accumulating in the intestine. This accumulation is a result of estradiol receptors present throughout the intestinal track (Zielonka et al., 2015).
In a study (Piotrowska et al., 2014), the influence of ZEN on colon microbiota diversity was assessed. Young gilts were fed with ZEN alone and ZEN in combination with DON (40 μg ZEN/kg BW/day and 12 μg DON/kg BW/day) for six weeks. ZEN had an adverse effect on mesophilic aerobic bacteria in a time-dependent manner. The combination of the two mycotoxins altered the microbiota’s metabolism, resulting in the formation of pro-carcinogenic substances.
In a later study (Cieplinska et al., 2019), ZEN exposure to young gilts resulted in differences on microbial counts between distal and proximal intestinal segments regarding microbiome diversity. In addition, ZEN boosted mold and yeast counts throughout the intestine. Similarly, changes in composition and biological functions have been reported in pigs challenged with a combination of ZEN and DON (Le Sciellour et al., 2020).
Immune system function is closely linked with the enteric nervous complex in the intestine. Intestine neuronal structure interacts with estrogen receptors present there. Administration of ZEN (10 μg/kg BW; 6 weeks) changed the activity of intramuscular nerve fibres (Gonkowski et al., 2015).

ZEN may also attribute to local inflammation in the intestine. Exposure of ZEN (40 μg/kg BW), DON (12 μg/kg BW) and a combination of both toxins increased the presence of black-brown granules in the lamina propria, indicating oxidative stress (Lewczuk et al., 2015).
Induction of oxidative stress in the intestine by ZEN was demonstrated in a recent study (Cheng et al., 2019). Piglets were fed with different levels of ZEN (0 to 1.5 ppm) for 35 days. ZEN induced oxidative stress on the jejunum segment via a specific signalling pathway (Keap1–Nrf2). High dietary levels of ZEN (1 ppm), induced oxidative stress in the small intestine. In addition, these levels reduced disaccharidase (enzymes such as lactase, sucrase, and maltase) activities, thus potentially affecting nutrient absorption (Liu et al., 2020).
ZEN and its metabolites (α-ZEL, β-ZEL) can affect intestinal epithelial integrity and functionality. In an in vitro study, all tested compounds demonstrated cytotoxicity. In addition, ZEN metabolites show a decrease on transepithelial electrical resistance by the end of experimental period (Marin et al., 2015).
Jia et al., (2020) showed the negative effect of ZEN on gut integrity in vivo. In piglets fed with ZEN (265 ppb) for three weeks, expression of claudin-4 (tight junction protein) was decreased. In addition, ZEN contributed to intestine inflammation. Furthermore, in combination with DON (1 ppm), ZEN increased pro-inflammatory cytokine levels and altered the cecum microbiota profile.
The connection between the generalized and reproductive effects of ZEN can be seen in fish larvae. ZEN induced developmental defects like pericardial edema, hyperemia, yolk sac edema, spine curvature and a heart rate reduction in zebrafish (Danio rario) .Neurotoxicty, DNA damage, cell apoptosis and structural histological changes were also reported in embryos of this model fish species. These, shed light, not only on ZEN’s toxic effects and mechanisms but also on the importance of the toxin expousre timing.
Similarly to other animals, the system and the stage most notibly influenced by ZEN are the reproductive system in breeding animals and early organ-system development in the offspring during embryonic/larval and juvenile life stages.
Conclusion
ZEN frequently occurrs in feedstuffs and is a major concern in animal production. Its negative effects in reproduction are well documented. Animal producers around the world are experiencing economic loses resulting from reproductive failure. Past and recent literature elucidates the negative effects of ZEN in immunity and gut health. Disruption of immunocompetence and gut health can pose an extra toll on animal welfare and farm profitability.
26 February 2021
Our experts are always available to help.
If you are an existing customer, please go directly to our Customer Portal.
We detected that you are visitng this page from United States. Therefore we are redirecting you to the localized version.
