-
Global/EN
- Global
- North America
- Latin America
In Brief
In this article, you’ll learn how ZEN can damage the reproductive potential of your pigs, signs and symptoms of problems in the herd and actions you can take to prevent or remedy them.
Reproductive performance is fundamental to the economic success of swine production operations, but zearalenone, a common mycotoxin, is a frequent cause of reproduction problems.
Reproductive performance is fundamental to the economic success of swine production operations. If reproductive problems occur, farms suffer tremendous losses due to a reduced number of piglets. Additionally, both determining the cause of a decline in reproductive performance and implementing corrective measures can incur high costs.
Zearalenone (ZEN) can severely impact reproductive performance in pigs due to its estrogenic effects (Zinedine et al., 2006). Hence, ZEN should always be considered as a differential diagnosis in case of reproductive failure. Understanding how ZEN works to undermine reproduction in your animals, being aware of what symptoms to look for and knowing how to prevent problems before they occur is critical to maximizing performance—and profit—in swine operations.
This means that feed should be monitored for ZEN and other relevant mycotoxins systematically. Having profound knowledge about toxin burden in the respective feed lot is the basis to review the implemented mycotoxin risk management strategy. If applicable, changes of dosage or used product can be performed based on the results.
Pigs are the livestock species most affected by the estrogenic effects of ZEN. ZEN binds to specific estrogen receptors and simulates the effects of this hormone (Osweiler, 2000). Subsequently the endocrine system is disrupted, which results in different outcomes in dependence of sex, age and reproductive status. The high susceptibility of swine to ZEN may be at least partly due to its metabolic pathways, which is driven to the more estrogenic metabolite α-ZEL (Biehl et al., 1993; Malekinejad et al. 2006). In fact, the first published reports of hyperestrogenism, retrospectively supposed to be caused by ZEN, were in swine (MacNutt et al., 1928; Pullar and Lerew, 1937; McErlean, 1952).
The European Commission (EC) identified the risk originating from ZEN and defined recommended thresholds for complementary and complete feeding stuffs (2006/576/EG). The differences of susceptibility of age groups are reflected by the guidance levels, which are 100 (gilts and piglets) and 250 ppb (sows and fatteners), respectively.
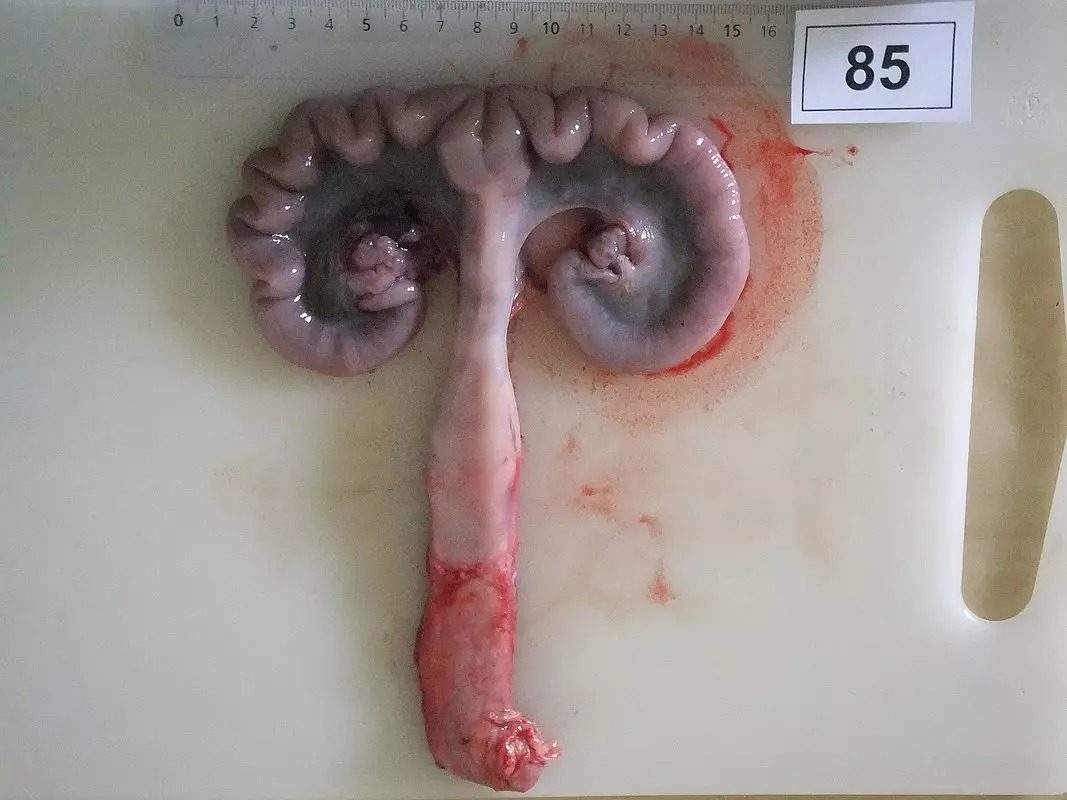
Figure 1a) Piglet was exposed to ZEN
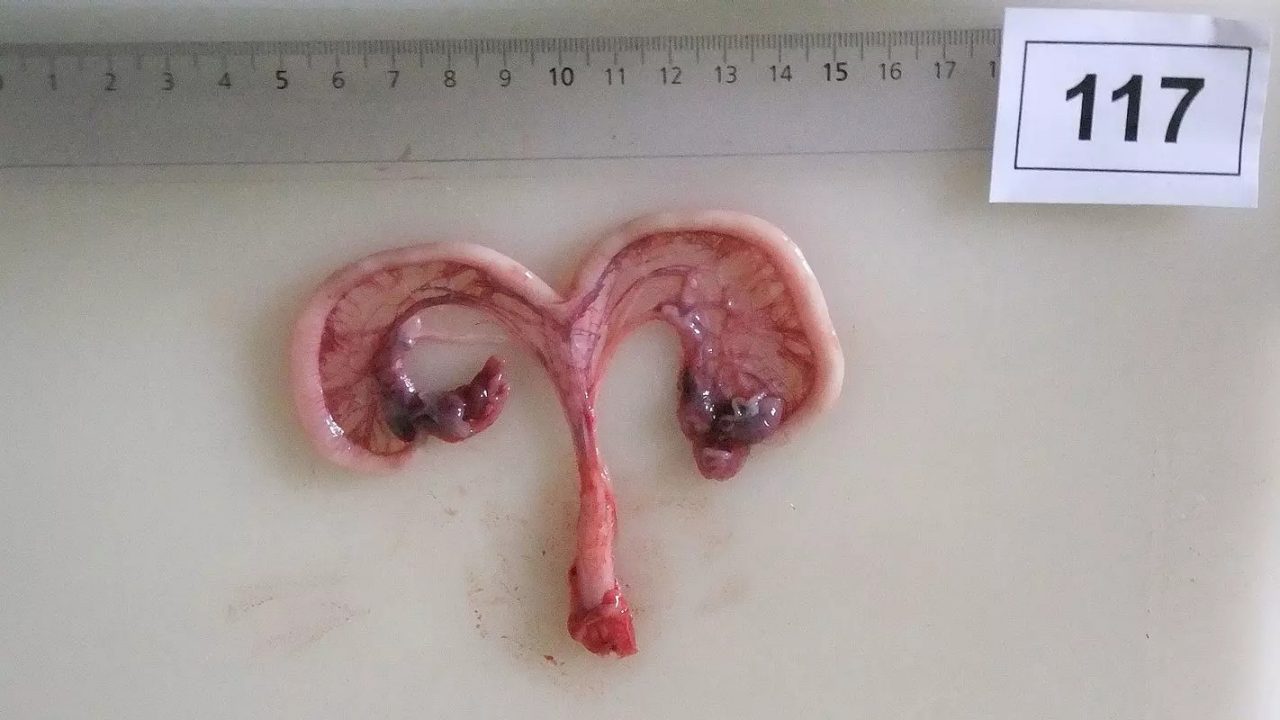
Figure 1b: Piglet received uncontaminated diet
Clinical presentation in female pigs is extremely variable depending on the stage of the reproductive cycle in which ZEN is ingested, dosage and duration of exposition (Osweiler, 2000). Especially, but not exclusively, the reproductive system of sexually immature gilts is affected by this estrogenic substance as well, with effects including:
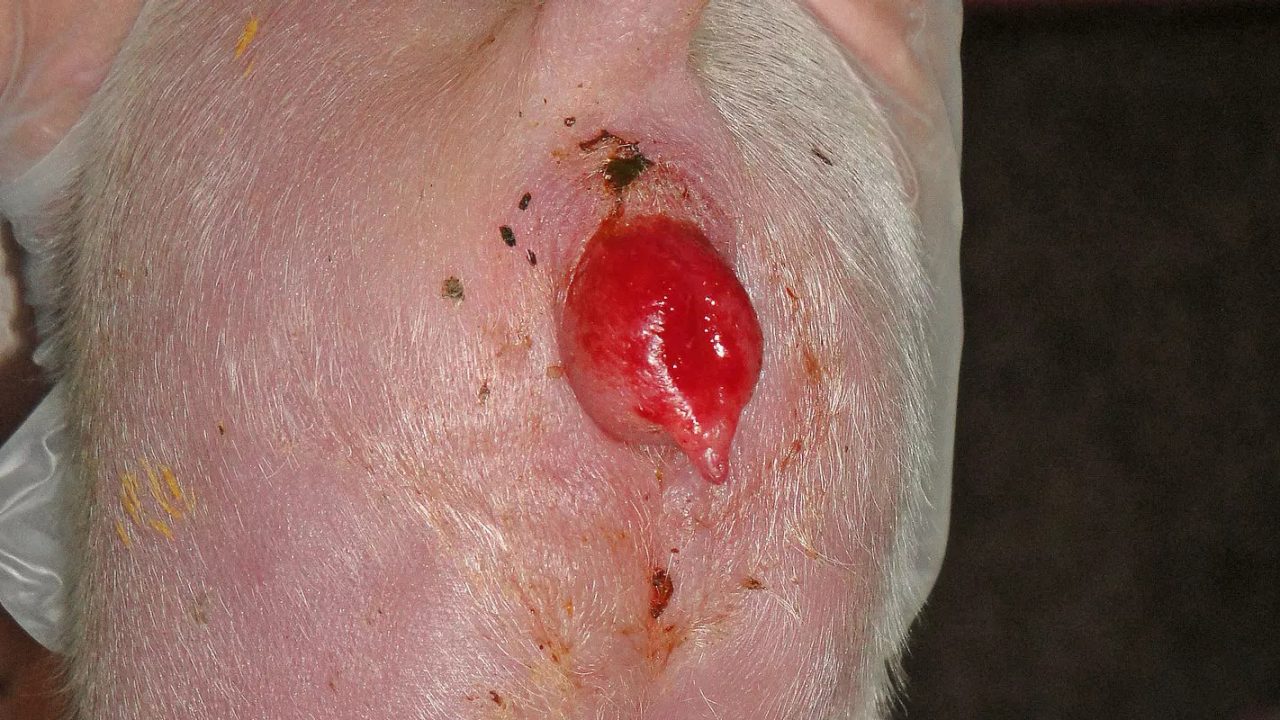
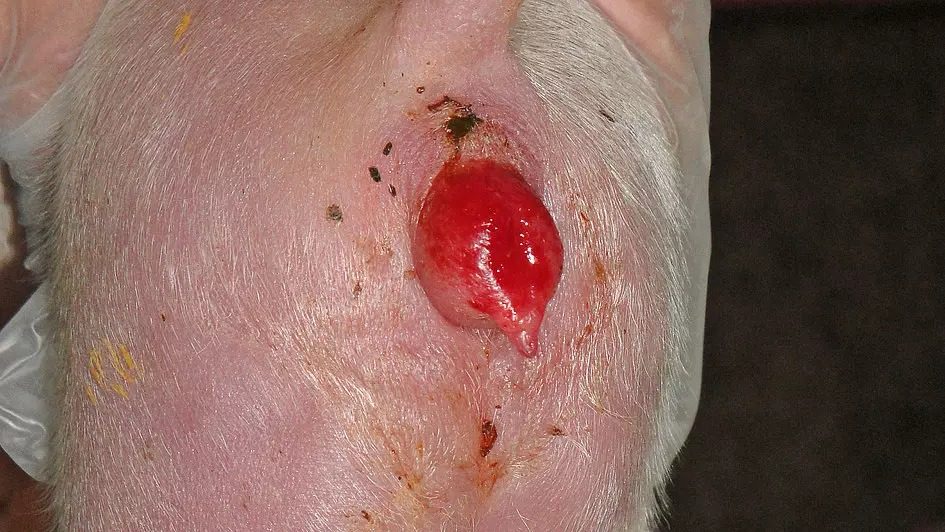
Figure 2: Juvenile hyperestrogenism of the offspring of sows exposed to ZEN
Copyright: Traunkreis Vet Clinic, Ried im Traunkreis
Detrimental effects of ZEN can also be seen in sexually mature gilts and sows. Clinical presentation depends on the phase of reproductive cycle, in which ZEN is ingested, dosage and duration of exposition, but includes:
In addition to ZEN’s impact on females, negative effects of ZEN on boars´ reproductive performance have been reported. Observable effects include:
The impact is possibly due to reduction of testosterone production by the hormone producing Leydig cells in the testicles (Yang et al., 2007).
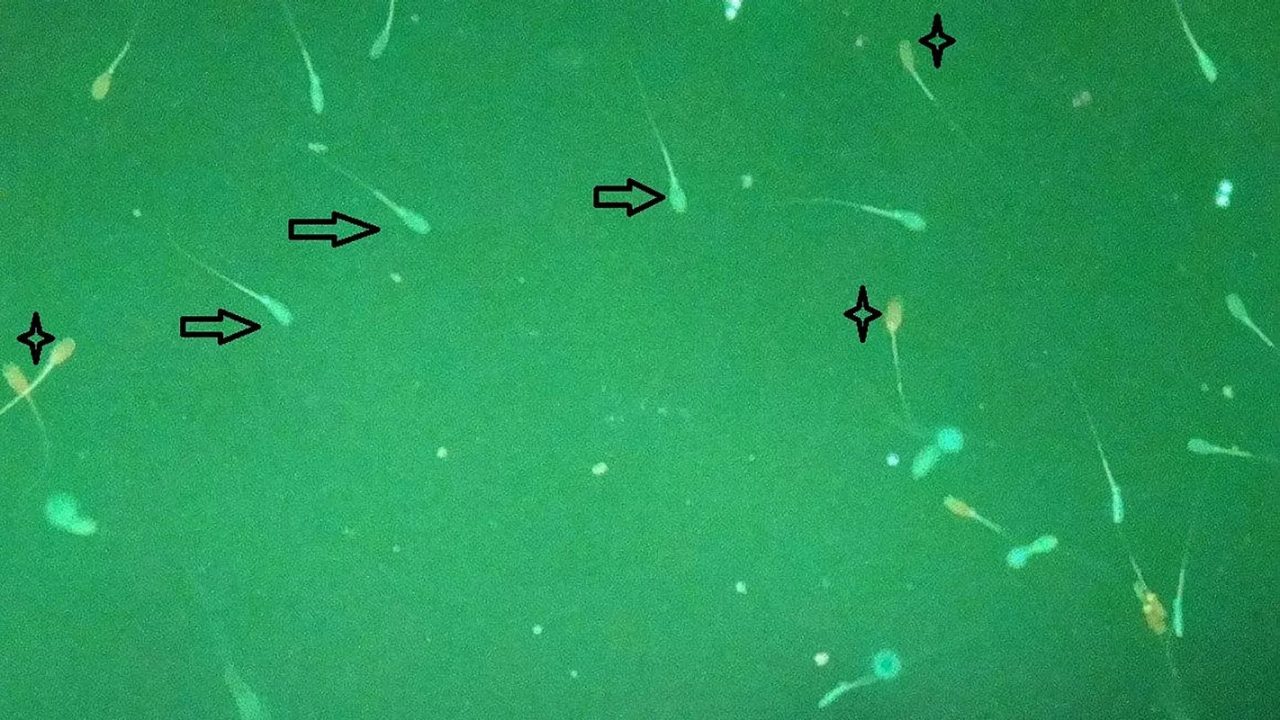
Figure 3: Spermatozoa exposed to ZEN: Differentiation based on fluorescence in alive (arrow) and dead (star sign) spermatozoa (Tassis et al., 2020)
In addition to observable effects in boars, direct effects of ZEN on spermatozoa could be proven in vitro (Figure 3), such as:
This may be mediated by binding of ZEN to estrogen receptors located on spermatozoa as they were found by Rago et al. (2007).
The dsm-firmenich World Mycotoxin Survey reveals that ZEN is one of the most prevalent well-known mycotoxins found worldwide, following Deoxynivalenol (DON) and Fumonisin (FUM) (Gruber-Dorninger et al., 2019). Within the last 10 years, prevalence of ZEN-positive corn kernel samples ranged between 45-55%. Average of concentration within positive samples was between 123 and 444 ppb in the respective period of time. Wheat can also significantly contribute to toxin burden within the feed. 32-52% of wheat samples have been identified as ZEN-positive within the last 10 years with average concentrations ranging from 55-102 ppb.
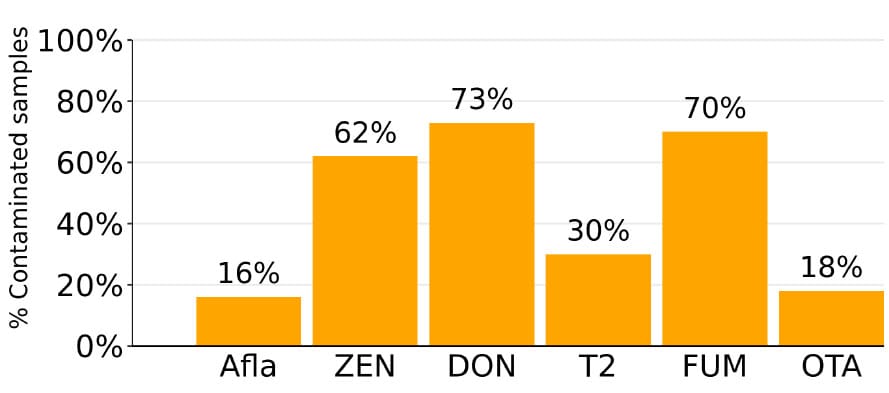
Figure 4: Prevalence of main mycotoxins in finished swine feed 2014-June 2023. Source: dsm-firmenich World Mycotoxin Survey
It is important to know that contamination levels below the regulatory limits can negatively affect the performance of pigs. This can be due to:
ZEN regularly co-occurs with other mycotoxins with potential impact on health and reproduction. Frequently, ZEN co-occurs with Deoxynivalenol (DON) as both toxins are produced by Fusarium spp. In contrast to ZEN, DON influences reproductive performance mainly indirectly for example by reduced feed intake (Tiemann and Dänicke, 2007). Recently it could be shown, that ZEN and DON reveal additive and synergistic effects on particular parameters, which are indicative for viability and velocity of spermatozoa (Tassis et al., 2020). As well ZEN as DON can inhibit maturation of pig oocyte maturation and therefor impair fertility. Co-occurrence probably leads to an additive effect (Malekinejad et al. 2007).
Fungi, plants and animals can modify mycotoxins. The resulting metabolites are not necessarily of less toxicity or estrogenicity than the parental substance. For example, the metabolite α-ZEL is known to be 60 times as estrogenic as ZEN (Lorenz et al., 2019). Some metabolites of mycotoxins may elude analytics and are therefore called masked mycotoxins (Berthiller et al., 2013). Masked mycotoxins lead to underestimation of toxin burden. It has been shown that during digestion in swine, toxic substances were set free from masked ZEN-metabolites (Gareis et al, 1990). Hence, if the different metabolites of ZEN are not covered by the applied diagnostic methods, overall toxin burden may be underestimated.
Underestimation of toxin burden can also be caused by a simple reason: mycotoxins are not evenly distributed in feed and therefor analyzed samples are not necessarily representative for the whole batch (Figure 5).

Figure 5: Schematic showing inhomogeneous distribution of mycotoxins (dark orange)
In swine production, health problems are often caused by multiple factors. It can be assumed that severity of mycotoxin-induced effects could be influenced by deficiencies regarding general health and immune status, herd management and stress exposure. Of course, the situations on individual farms are difficult to simulate in a scientific model and therefor the effects of different factors are difficult to assess. However, experience and a recent case report indicate significance of low levels of ZEN and its metabolites under practical conditions (Hennig-Pauka et al. 2018).
Besides the Fusarium-derived ZEN, diets can also contain phytoestrogens. These substances occur naturally in soybeans, clover and other legumes (Nikov et al., 2000). In cases of hyperestrogenism in animals, which cannot be explained by exposure to ZEN, rations should be controlled for phytoestrogens.
Conclusion
ZEN is a frequently occurring contaminant, and its effect on reproductive performance can be severe. Hence, a proper mycotoxin risk management strategy should be implemented on every farm to avoid negative impacts on swine health and performance. Be aware, that impairment of reproduction is the predominant outcome of ZEN ingestion, but other organ systems can be affected as well.
Tips
Act preventively and test feed regularly.
ZEN is one of the most prevalent mycotoxins worldwide.
Do not underestimate ZEN.
Even concentrations below regulatory limits can severely effect reproduction.
Reduce pigs’ toxin burden.
Lower levels of toxins can help your animals reach their full potential.
References
Alm H., Brussow, K.P., Torner, H., Vanselow, J., Tomek, W., Danicke, S., Tiemann, U. (2006). Influence of Fusarium-toxin contaminated feed on initial quality and meiotic competence of gilt oocytes. Reprod Toxicol 22: 44-50.
Benzoni, E., Minervini, F., Giannoccaro, A., Fornelli, F., Vigo, D., Visconti, A. (2008). Influence of in vitro exposure to mycotoxin zearalenone and its derivatives on swine sperm quality. Reproductive toxicology, 25(4), 461-467.
Berger, T., Esbenshade, K.L., Diekman, M.A., Hoagland, T., Tuite, J. (1981). Influence of prepubertal consumption of zearalenone on sexual development of boars. J Anim Sci 53: 1559-1564.
Berthiller, F., Crews, C., Dall'Asta, C., Saeger, S. D., Haesaert, G., Karlovsky, P., Oswald, I.P., Seefelder, W., Speijers, G., Stroka, J. (2013). Masked mycotoxins: A review. Molecular nutrition & food research, 57(1), 165-186.
Biehl, M. L., Prelusky, D. B., Koritz, G. D., Hartin, K. E., Buck, W. B., Trenholm, H. L. (1993). Biliary excretion and enterohepatic cycling of zearalenone in immature pigs. Toxicology and applied pharmacology, 121(1), 152-159.
Bielas, W., Niżański, W., Nicpoń, J., Nicpoń, J. E., Partyka, A., Mordak, R., Nowak. M., Ciaputa, R. (2017). Effect of zearalenone on circulating testosterone concentration, testicular and epididymal morphology and epididymal sperm characteristics in wild boars. Theriogenology, 102, 59-66.
Chang, K., Kurtz, H.J., Mirocha, C.J. (1979). Effects of the mycotoxin zearalenone on swine reproduction. Am J Vet Res 40: 1260-1267.
Chen, X.X., Yang, C.W., Huang, L.B., Niu, Q.S., Jiang, S.Z., Chi, F. (2015). Zearalenone altered the serum hormones, morphologic and apoptotic measurements of genital organs in post-weaning gilts. Asian-Australas J Anim Sci 28: 171-179.
Christensen, C.M., Mirocha, C.J., Nelson, G.H., Quast, J.F. (1972). Effect on young swine of consumption of rations containing corn invaded by Fusarium roseum. Appl Microbiol 23: 202.
Edwards, S., Cantley, T.C., Day, B.N. (1987). The effects of zearalenone on reproduction in swine. II. The effect on puberty attainment and postweaning rebreeding performance. Theriogenology 28: 51-58.
Etienne, M., Jemmali, M. (1982). Effects of zearalenone (F2) on estrous activity and reproduction in gilts. J Anim Sci 55: 1-10.
Gajecka, M., Rybarczyk, L., Zwierzchowski, W., Jakimiuk, E., Zielonka, L., Obremski, K., Gajecki, M. (2011). The effect of experimental, long-term exposure to low-dose zearalenone mycotoxicosis on the histological condition of ovaries in sexually immature gilts. Theriogenology 75: 1085-1094.
Gareis, M., Bauer, J., Thiem, J., Plank, G., Grabley, S., & Gedek, B. (1990). Cleavage of zearalenone‐glycoside, a “masked” mycotoxin, during digestion in swine. Journal of Veterinary Medicine, Series B, 37(1‐10), 236-240.
Grenier, B., Hackl, M., Skalicky, S., Thamhesl, M., Moll, W.D., Berrios, R., Schatzmayr, G., Nagl, V. (2019). MicroRNAs in porcine uterus and serum are affected by zearalenone and represent a new target for mycotoxin biomarker discovery. Sci Rep 9: 9408.
Gruber-Dorninger, C., Jenkins, T., Schatzmayr, G. (2019). Global mycotoxin occurrence in feed: A ten-year survey. Toxins, 11(7), 375.
Hennig-Pauka, I., Koch, F. J., Schaumberger, S., Woechtl, B., Novak, J., Sulyok, M., & Nagl, V. (2018). Current challenges in the diagnosis of zearalenone toxicosis as illustrated by a field case of hyperestrogenism in suckling piglets. Porcine health management, 4(1), 18.
Kordić, B., Pribićević, S., Muntanola-Cvetković, M., Nikolić, P., & Nikolić, B. (1992). Experimental study of the effects of known quantities of zearalenone on swine reproduction. Journal of Environmental Pathology, Toxicology and Oncology: Official Organ of the International Society for Environmental Toxicology and Cancer, 11(2), 53-55.
Liu, J., & Applegate, T. (2020). Zearalenone (ZEN) in Livestock and Poultry: Dose, Toxicokinetics, Toxicity and Estrogenicity. Toxins, 12(6), 377.
Long, G.G., Turek, J., Diekman, M.A., Scheidt, A.B. (1992). Effect of zearalenone on days 7 to 10 post-mating on blastocyst development and endometrial morphology in sows. Vet Pathol 29: 60-67.
Lorenz, N., Dänicke, S., Edler, L., Gottschalk, C., Lassek, E., Marko, D., Rychlik, M., Mally, A. (2019). A critical evaluation of health risk assessment of modified mycotoxins with a special focus on zearalenone. Mycotoxin research, 35(1), 27-46.
Malekinejad, H., Schoevers, E.J., Daemen, I.J., Zijlstra, C., Colenbrander, B., Fink-Gremmels, J., Roelen, B.A. (2007). Exposure of oocytes to the Fusarium toxins zearalenone and deoxynivalenol causes aneuploidy and abnormal embryo development in pigs. Biol Reprod 77: 840-847.
Malekinejad, H., Maas-Bakker, R., Fink-Gremmels, J. (2006). Species differences in the hepatic biotransformation of zearalenone. The Veterinary Journal, 172(1), 96-102.
McErlean, B. A. (1952) Vulvovaginitis of Swine. Vet. Record, 64: 539-540.
McNutt, S. H., Purwin, P., & Murray, C. (1928). Vulvovaginitis in swine. J. Am. Vet. Med. Assoc, 73(484), 1928.
Nikov, G. N., Hopkins, N. E., Boue, S., & Alworth, W. L. (2000). Interactions of dietary estrogens with human estrogen receptors and the effect on estrogen receptor-estrogen response element complex formation. Environmental Health Perspectives, 108(9), 867-872.
Obremski, K. (2014). The effect of in vivo exposure to zearalenone on cytokine secretion by Th1 and Th2 lymphocytes in porcine Peyer's patches after in vitro stimulation with LPS. Pol J Vet Sci 17: 625-632.
Oliver, W.T., Miles, J.R., Diaz, D.E., Dibner, J.J., Rottinghaus, G.E., Harrell, R.J. (2012). Zearalenone enhances reproductive tract development, but does not alter skeletal muscle signaling in prepubertal gilts. Anim Feed Sci Tech 174 79-85.
Osweiler, G. D. (2000). Mycotoxins: Contemporary issues of food animal health and productivity. Veterinary Clinics: Food Animal Practice, 16(3), 511-530.
Pullar, E. M., Lerew, W. M. (1937). Vulvovaginitis of swine. Australian Veterinary Journal, 13(1), 28-31.
Rago, V., Aquila, S., Panza, R., Carpino, A. (2007). Cytochrome P450arom, androgen and estrogen receptors in pig sperm. Reproductive Biology and Endocrinology, 5(1), 1-6.
Tassis, P.D., Tsakmakidis, I.A., Nagl, V., Reisinger, N., Tzika, E., Gruber-Dorninger, C., Michos, I., Mittas, N., Basioura, A., Schatzmayr, D. (2020). Individual and combined in vitro effects of deoxynivalenol and zearalenone on boar semen. Toxins (Basel) 12: 495.
Teixeira, L.C., Montiani-Ferreira, F., Locatelli-Dittrich, R., Santin, E., Alberton, G.C. (2011). Effects of zearalenone in prepubertal gilts. Pesq Vet Bras 31: 656-662.
Tiemann, U., Dänicke, S. (2007). In vivo and in vitro effects of the mycotoxins zearalenone and deoxynivalenol on different non-reproductive and reproductive organs in female pigs: a review. Food additives and contaminants, 24(3), 306-314.
Tsakmakidis, I. A., Lymberopoulos, A. G., Alexopoulos, C., Boscos, C. M., & Kyriakis, S. C. (2006). In vitro effect of zearalenone and α‐zearalenol on boar sperm characteristics and acrosome reaction. Reproduction in domestic animals, 41(5), 394-401.
Yang, L.J., Zhou, M, Huang L.B., Yang, W.R., Yang, Z.B., Jiang, S.Z., Ge, J.S. (2018). Zearalenone-promoted follicle growth through modulation of Wnt-1/beta-catenin signaling pathway and expression of estrogen receptor genes in ovaries of postweaning piglets. J Agric Food Chem 66: 7899-7906.
Yang, J. Y., Wang, G. X., Liu, J. L., Fan, J. J., & Cui, S. (2007). Toxic effects of zearalenone and its derivatives α-zearalenol on male reproductive system in mice. Reproductive Toxicology, 24(3-4), 381-387.
Young, L.G., King, G.J. (1986). Low concentrations of zearalenone in diets of mature gilts. J Anim Sci 63: 1191-1196.
Young, L.G., Ping, H., King, G.J. (1990). Effects of feeding zearalenone to sows on rebreeding and pregnancy. J Anim Sci 68: 15-20.
Zhou, M., Yang, L.J., Yang, W.R., Huang, L.B., Zhou, X.M., Jiang, S.Z., Yang, Z.B. (2018) Effects of zearalenone on the localization and expression of the growth hormone receptor gene in the uteri of post-weaning piglets. Asian-Australas J Anim Sci 31: 32-39.
Zinedine, A., Soriano, J. M., Molto, J. C., & Manes, J. (2007). Review on the toxicity, occurrence, metabolism, detoxification, regulations and intake of zearalenone: an oestrogenic mycotoxin. Food and chemical toxicology, 45(1), 1-18.
Zwierzchowski W., Przybylowicz, M., Obremski, K., Zielonka, L., Skorska-Wyszynska, E., Gajecka, M., Polak, M., Jakimiuk, E., Jana, B., Rybarczyk, L., Gajecki, M. (2005). Level of zearalenone in blood serum and lesions in ovarian follicles of sexually immature gilts in the course of zearalenone micotoxicosis. Pol J Vet Sci 8: 209-218.
02 August 2023
Our experts are always available to help.
If you are an existing customer, please go directly to our Customer Portal.
We detected that you are visitng this page from United States. Therefore we are redirecting you to the localized version.
