
PoultryStar®
PoultryStar® is a well-defined, multi-species poultry probiotic plus prebiotic (synbiotic) that promotes a beneficial gut microflora and improves the results of broilers, layers and breeders.
Poultry coccidiosis is one of the most common diseases in the world. It generates economic losses due to mortality, reduced body weight plus the expenses related to preventive and therapeutic control, while also predisposing birds to necrotic enteritis. Worldwide yearly losses have been estimated at more than US$3 billion due to coccidiosis in chickens and other avian species.
Coccidiosis Control
Enhancing your coccidiosis control program to reduce the reliance on anticoccidials as well as boosting the efficacy of coccidiosis vaccines.
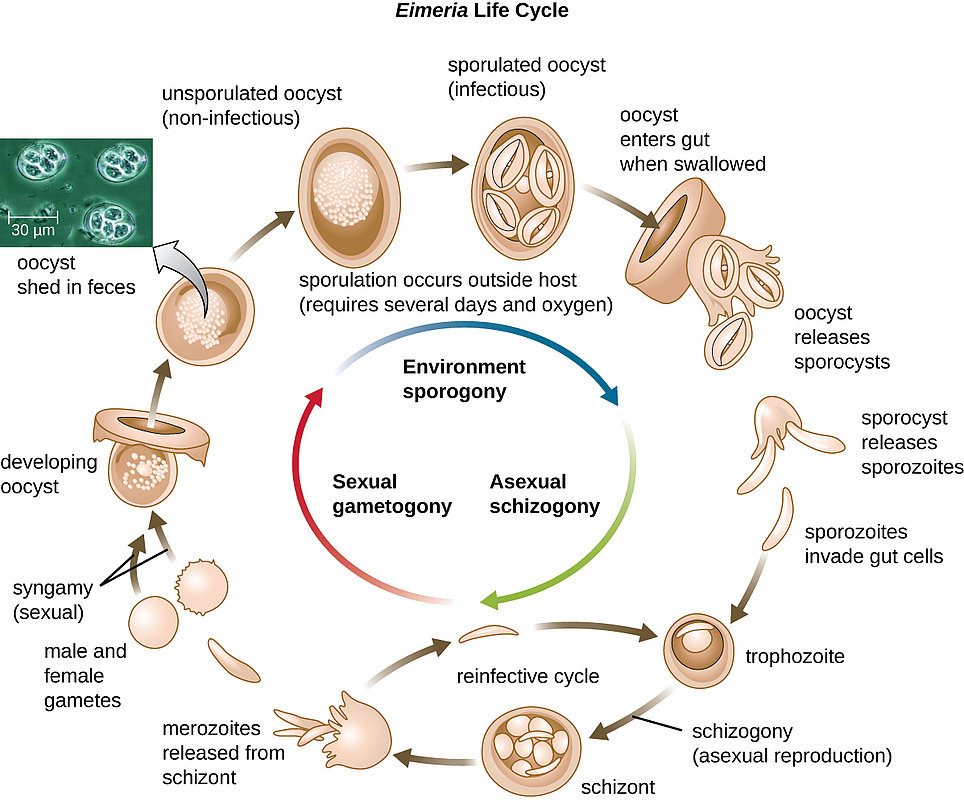
Figure 1: The Eimeria life cycle. Eimeria cause coccidiosis in chickens and other avian species.
Coccidiosis is caused by protozoan parasites of the genus Eimeria. These are obligated intracellular parasites with complex life cycles including sexual and asexual stages. In poultry, Eimeria affect the intestine making it prone to other diseases (necrotic enteritis) and reducing the ability of this organ to absorb nutrients.
Modern poultry production practices facilitate the distribution of this disease within poultry houses. Between poultry houses, the disease is transmitted by mechanical carriers such as insects and wild birds. While Eimeria eggs (oocysts) can be mechanically transported by wild birds, these parasites are host-specific and thus wild birds do not serve as a biological reservoir.
The life cycle of Eimeria starts with the ingestion of mature oocysts. Each infective oocyst is formed by four sporocysts and in turn each sporocyst contains two sporozoites. Bile salts and chymotrypsin stimulate the release of the sporozoites from the oocyst. Once freed, the sporozoites invade intestinal cells beginning the asexual development stage called schizogony. After a variable number of asexual cycles, gametes are formed and the sexual stage of development begins (gamogony). The sexual phase terminates with the production and release of oocysts into the intestinal lumen. Once in the environment, oocysts must sporulate to become infective. Sporulation process usually takes from 2 to 3 days depending on environmental conditions (Waldenstedt et al., 2001).
In general, good natural immunity is generated after Eimeria infections in poultry and for this reason coccidiosis is usually a disease that affects young animals. However, the achieved immunity is specific for each of the species of Eimeria and it is not cross-protective between species (with the exception of some cross protection between E. maxima and E. brunetti). Even more, cross-protection among strains of the same species is often partial (Long, 1974), which is a practical concern for the selection and use of live vaccines against Eimeria in different geographical locations.
Characteristics of lesions will depend on the species of Eimeria affecting the intestine. Important diagnostic data to determine which Eimeria species is affecting a particular flock include:
Most common Eimeria species for poultry:
This species of Eimeria is widely distributed in commercial poultry productions. Mortality may result from heavy infections but often a reduction in weight gain and skin pigmentation (due to a reduced intestinal absorption) are the most predominant features of Eimeriaacervulina.
Lesions of Eimeria acervulina locate in the small intestine. In light infections the lesions usually concentrate in the duodenum but in heavy infections the lesions can extend beyond the duodenum into the rest of the small intestine.
Lesions can be observed from the serosal surface of the intestine as white plaques that tend to arrange forming transversal striations of the duodenum. The intestinal mucosa can be thickened and can be covered by clear fluid.
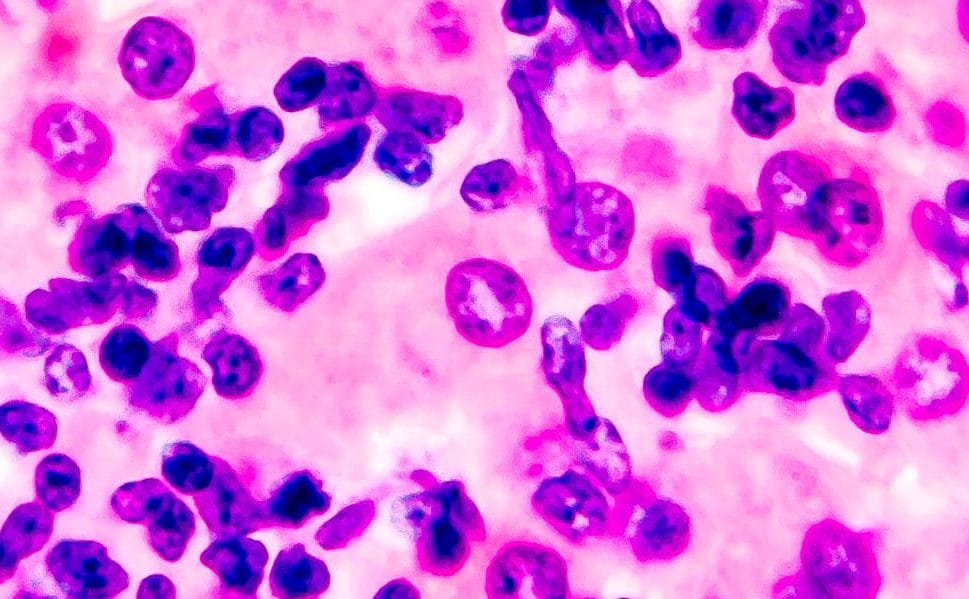
Named after the large size of their oocysts, Eimeriamaxima is moderately pathogenic. It causes losses in body weight and decreases skin pigmentation due to the reduced absorption of pigments due to intestinal damage.
Birds can be emaciated due to reduced feed intake coupled with poor nutrient absorption. Some mortality can be seen in severe cases.
Eimeriamaxima preferentially colonizes the medium portion of the small intestine but in severe cases lesions can totally cover the small intestine. The lumen of the intestine may contain orange mucus and blood, and in heavy infections the mucosa can be seriously damaged.
This species of Eimeria can induce poor feed conversion, loss in weight gain, and moderate mortality in severe infections. In severe cases feces can be stained with blood.
Eimeria brunetti locates preferentially in the ileum but in severe cases the lesions extend towards the large intestine and the upper portions of the small intestine.
Light infections are characterized by thickening of the intestinal mucosa and presence of petechiae in the lower part of the small intestine.
In severe cases, villi are almost completely denuded and the mucosa of the small intestine can be extremely damaged and necrotic.
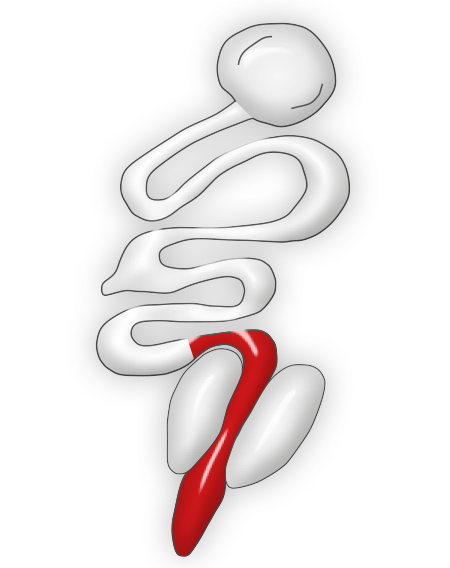
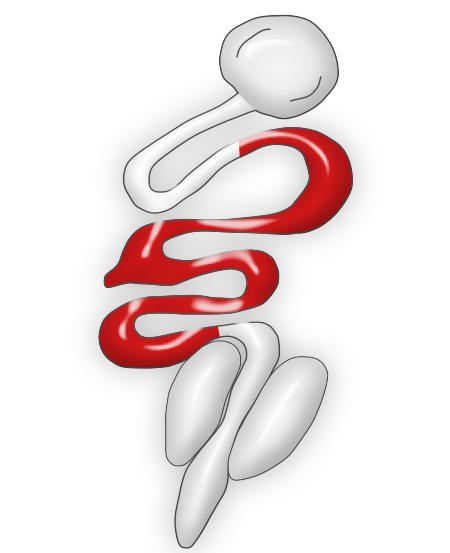
This species of Eimeria is highly pathogenic in chickens and it is often seen in birds from 9 to 14 weeks of age (McDouglas and Reid, 1991). Mortality, severe weight losses, and feces with blood and mucus are frequent findings.
Eimeria necatrix produces lesions in the medium portion of the small intestine. The intestine is usually dilated and constricted in some locations with a ballooned appearance, and the lumen often contains blood, mucosal debris, and fluid. Lesions can be seen from the serosal surface like white and dark dots that are usually described to have a "salt and pepper" appearance.
This species of Eimeria is highly pathogenic for chickens. E. tenella affects mostly the ceca causing spectacular lesions. High mortality, severe weight losses, and feces stained with blood are frequent findings. Severe weight loss is usual and in comparison to healthy flocks, the affected flocks usually cannot compensate for the weight loss after the disease has been controlled.
E. tenella produces hemorrhage in both ceca which is accompanied by the presence of white dots (schizonts and oocysts) that can be seen from the serosal surface. Eimeria tenella penetrates deep in the intestinal tissue producing heavy damage in the mucosa and muscular layer. The cecal lumen is filled with coagulated blood and necrotic mucosal debris.

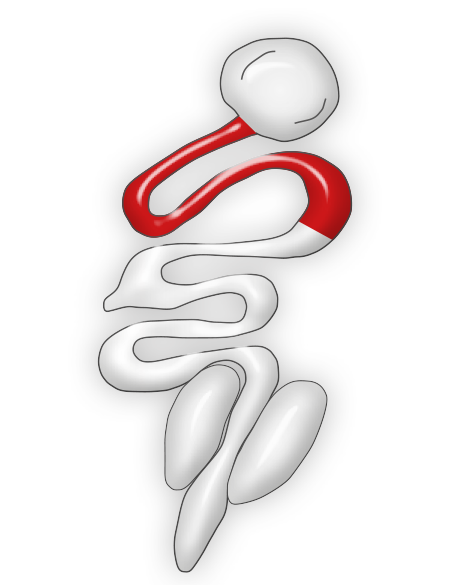
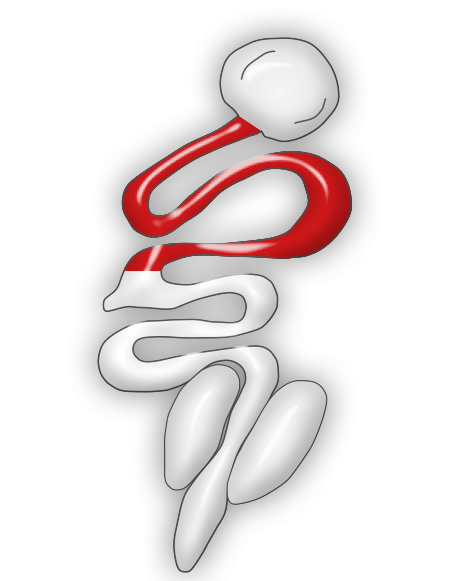
For turkeys this species of Eimeria is very pathogenic. It may produce mortality and important weight loss. Feces may be stained with blood and may contain mucus.
Eimeria adenoeides normally affects the ceca but lesions could extend to the surrounding portions of the intestine. Ceca are found edematous, distended, and filled with material that hardens into a caseous core after several days of the initial infection.
Eimeria meleagrimitis affects the upper portion of the small intestine but may also affect lower regions in severe infections. Less pathogenic than E. adenoeides, E. meleagrimitis can produce dehydration, weight losses, and some mortality in heavy infections.
The duodenum may appear congested and its lumen is filled with mucus and fluid. Hemorrhage is rare but possible.
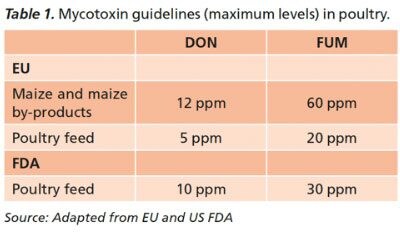
Mycotoxins in poultry feeds, even at permitted levels, aggravate the incidence and severity of coccidiosis in chickens, ducks and turkeys. They can facilitate Eimeria colonization in the gut, and poultry producers looking to reduce the impact of coccidiosis on their flocks would do well to monitor feed for contamination.
Deoxynivalenol and fumonisins are known to interfere with several vital functions of cells, and disrupt intestinal cells that act as a barrier between pathogens and the birds’ bodies. These disrupted intestinal cell components can be used as a growth substrate for pathogens such as Eimeria, Clostridium and Escherischia coli.
In this way, these mycotoxins play a main role as ‘gate openers’ favoring pathogen colonization throughout the host.
Lesion scoring in the cecum was 1.33 for the mycotoxin groups versus 0.42 for birds fed control diets (Figure 1). The number of oocysts found in the jejunum was three times higher for birds fed mycotoxins versus control. Similarly, the number of oocysts recorded in feces was 29 percent higher for the deoxynivalenol + fumonisins group and 46 percent higher for the fumonisins group.
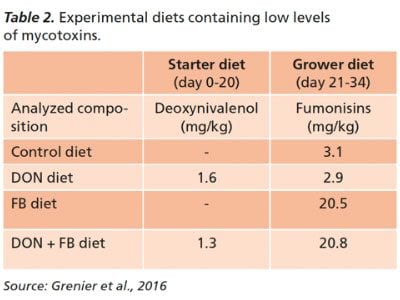
Even at concentrations allowed under European and US guidelines for mycotoxins (Table 1), deoxynivalenol, fumonisins and a combination of the two can worsen the incidence and severity of coccidiosis in challenged poultry, studies have shown.
Chickens fed mycotoxin-contaminated feed at levels below regulatory guidelines in starter and grower diets (Table 2) displayed considerably higher lesion scores, higher numbers of oocysts in both the jejunum and in excreta, and higher lymphocyte (white blood cell) counts.
In poultry production several antimicrobials or antiprotozoals have been used for decades to treat and prevent coccidiosis. Depending on the type of poultry production, the approaches for an effective control of coccidiosis are different.
Methods of coccidiosis prevention or treatment:
Due to the short life cycle of a broiler the coccidiosis preventive program used usually aims for eliminating Eimeria completely from the gut by using coccidicides that kill the parasites. This results in optimal condition of the gastrointestinal tract, improving body weight, and reducing feed conversion (McDougald and Reid, 1991).
In breeders and layers a different approach is usually needed. Due to the relatively long life cycle of these birds, development of protective immunity is desired. For this purpose a minimal degree of exposure to Eimeria is allowed.
To achieve this objective, coccidiostats are used to arrest the development of the parasites at different stages of development allowing for a good balance between intestinal damage and appropriate exposure for immunity development. Of course, once the coccidiostats are withdrawn from the diet, the infecting parasites may resume their life cycle producing the clinical manifestations of the disease (McDougald and Reid, 1991).
Regardless of the drugs selected for controlling and treating coccidiosis, it is important to consider that Eimeria parasites do develop drug resistance. The resistance is greatly enhanced if the same family of antimicrobials is used for a long time within a defined area. Selective pressure will favor the few parasites within a population that are resistant, and within few rearing cycles the initial parasites would increase their population size to numbers able to induce clinical disease in a flock.
A common practice to partially solve this problem is to use anti-coccidial ‘shuttle’ programs that rotate through different periods of the bird's life. This method has a good chance of eliminating the parasites that demonstrated resistance to a single antimicrobial. A variation of the same principle consists on changing coccidiostats between flocks (Chapman, 2001).
A great proportion of the damage induced in coccidiosis is attributable to body weight losses. Part of the body weight losses can be explained due to suboptimal nutrient absorption in the intestine derived from epithelia destruction during and after Eimeria infection.
Another component of the body weight losses may be due to excessive inflammation developing in the gastrointestinal tract. Experimental models of bacterial infections have demonstrated that 41% of body weight losses are caused by lipopolysaccharide-induced inflammation. In this model, the use of PoultryStar® alleviated 17% of the growth depression probably by decreasing the amount of nutrients directed to the inflammatory process and thus increasing resources directed to increase body weight (Jiang et al., 2009).
The use of live vaccines is also common in broiler and layer breeders. The strategy with vaccination is to either use precocious strains that undergo only a few replication cycles in the avian intestine before shedding oocysts or to use attenuated strains and controlled dosing inducing less damage to the intestinal tract than a field strain (McDougald and Reid, 1991).
Vaccination can induce some decreases in body weight when compared to non-vaccinated groups using anticoccidial drugs (Chapman et al., 2002). In fact, a trial of probiotics administered with an Eimeria vaccine showed that birds receiving PoultryStar® had higher body weight than the birds that received the vaccine alone; in addition, the probiotic treatment raised the body weight to a level not different from the control group that used ionophores to control Eimeria (Klein et al., 2009).
Chapman, H.D. 2001. Use of anticoccidial drugs in broiler chickens in the USA: analysis for the years 1995 to 1999. Poult. Sci. 80:572-580.
Chapman, H.D., T.E. Cherry, H.D. Danforth, G. Richards, M.W. Shirley, andR.B. Williams. 2002. Sustainable coccidiosis control in poultry production: The role of live vaccines. Int. J. Parasitol. 32:617-629.
Dalloul, R.A., H.S. Lillehoj, T.A. Shellem, and J.A. Doerr. 2003. Enhanced mucosal immunity against Eimeria acervulina in broilers fed a Lactobacillus-based probiotic. Poult. Sci. 82:62-66.
Jiang Z., G. Schatzmayr, M. Mohnl, and T.J. Applegate. 2009. Net effect of an acute phase response: Alleviation with probiotic supplementation. PSA abstracts, North Carolina, USA.
Jang, S.I., M. Jun, H.S. Lillehoj, R.A. Dalloul, I. Kong, S. Kim, and W. Min. 2006. Anticoccidial effect of green tea-based diets against Eimeria maxima. Vet. Parasit. 144:172-175.
Klein, A., J. Lee, M. Farnell, L. Oden, S. Pohl, K. Stringfellow, M. Mohnl, R. Beltran, G. Schatzmayr, S. Fitz-Coy, C. Broussard, and D. Caldwell. 2009. Effects of probiotic administration during coccidiosis vaccination on performance in broilers exposed to field strain Eimeria: Comparison to monensin administration. PSA abstracts, North Carolina, USA.
Lee, S.H., H.S. Lillehoj, R.A. Dalloul, D.W. Park, Y.H. Hong, and JJ. Lin. 2007. Influence of Pediococcus-based probiotics on Coccidiosis in broiler chickens. Poult. Sci. 86:63-66.
Long, P.L. 1974. Experimental infection of chickens with two species of Eimeria isolated from the Malaysian jungle fowl. Parasitology 69:337-347.
McDougald, L.R. and W.M. Reid. 1991. Coccidiosis. In: Diseases of Poultry, ninth edition. Iowa State Press, Iowa.
Waldenstedt, L., K. El winger, A. Lunden, P. Thebo, and A. Uggla. 2001. Sporulation of Eimeria maxima oocysts in litter with different moisture content. Poult. Sci. 80:1412-1415.
Williams, R.B. 1999. A compartmentalized model for the estimation of the cost of coccidiosis in the world's chicken production industry. Int. J. Parasitol. 29:1209-1229.
PoultryStar® is a well-defined, multi-species poultry probiotic plus prebiotic (synbiotic) that promotes a beneficial gut microflora and improves the results of broilers, layers and breeders.
Symphiome™ is a complex glycan mixture technically defined as a Precision Biotic*. A first-of-its-kind microbiome metabolic modulator precisely designed to harness the power of the microbiome by modulating specific and highly conserved functional pathways to enhance the nutritional health and performance of the birds.
Harness the power of data to get a deeper understanding of the overall health, productivity and welfare of your animals.
Get in touch with a dsm-firmenich Animal Nutrition & Health specialist or find contacts around the world to suit your needs.
At dsm-firmenich, we love to connect with you.
Follow us on any of the channels below.





