
Biomin® BioStabil
Biomin® BioStabil is a formulation of strategically selected Lactic Acid Bacteria for optimal haylage, silage and forage preservation. Biomin® BioStabil preserves the energy in your silage.
Ruminal acidosis is a bovine metabolic disease that affects feedlot as well as dairy cattle. Acidosis in cattle is usually associated with the ingestion of large amounts of highly fermentable, carbohydrate-rich feeds, which result in the excessive production and accumulation of acids in the rumen.
Ruminal acidosis can be present in different forms, reaching from peracute life-threatening forms to chronic illness, which is difficult to detect (Oetzel, 2003).
The difference between acute and subacute forms are that during acute ruminal acidosis the pH depression is more severe (Oetzel et al., 1999), and clinical signs more prominent (Kleen et al., 2003). Acute rumen acidosis is common in feedlots, whereas SARA is more common on dairy farms (Krause and Otzel, 2006). Contrary to acute acidosis, in SARA the pH depression is apparently due to the total accumulation of volatile fatty acids alone and is not due to lactic acid accumulation (Krause and Otzel, 2006).

Subacute ruminal acidosis is the most important nutritional disease in dairy cattle since it can negatively impact the dairy industry by decreasing dry matter intake, milk production, profitability, and increasing culling rate and death loss (McCann et al., 2016).
The current definition of SARA is based on a time period during which the rumen pH is below a certain threshold. Although, there is no general agreement on the pH threshold for SARA, the two main definitions indicate 5.24 hours below 5.8 (Zebeli et al., 2008) and 3 hours below 5.6 (Plaizier at al. 2008).
Dr. Christina Gruber, Scientist at dsm-firmenich, provides an overview of rumen acidosis and a rumen simulation model used in the lab to find effective acidosis prevention strategies.
Among the scientific community, the most common diagnostic techniques are based on rumen pH determination. The most widespread methods are listed below:
SARA occurs when ruminal buffering is not adequate to contrast the volatile fatty acids (VFAs) production. This may be due to different reasons:
Cows are more at risk to develop SARA in the following circumstances:
Prevalence of SARA increases as cows consume more total dry matter and as cows consume diets containing higher proportion of grain. However, in intensive dairy farming, the problem is technically unavoidable. Group feeding and strong variability among individual cows in terms of rumen microbiome, are the main reasons of failure (Figure 1)
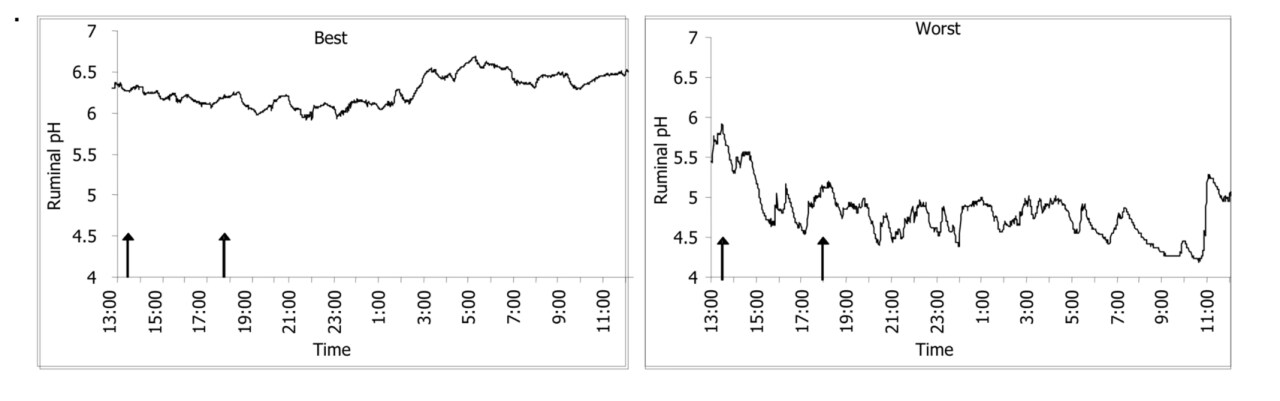
Figure 1. Ruminal pH measured 5 days after calving in two cows (best and worst-case acidosis cows) fed the same lactation diet (Penner, Beauchemin and Mutsvangwa, unpublished data)
| Prevalence | Country | Reference |
|---|---|---|
| 19% early lactation 26% mid-lactation | US (15 farms) | Garrett et al. 1997 |
| 20.1% early and peak lactation | US (14 farms) | Oetzel et al. 1999 |
| 13.8% overall | Netherlands | Kleen et al. 2009 |
| 11% grazing cows | Ireland | O'Grady et al. 2008 |
| 33% early lactation | Italy | Morgante et al. 2007 |
| 11% early lactation 18% mid-lactation | Germany/Netherlands | Kleen et al. 2004 |
| 20% overall | Germany | Kleen et al. 2013 |
| 14% overall | Poland | Stefanska et al. 2016 |
Table 1. SARA prevalence in dairy herds reported by different authors.
SARA prevalence ranges between 11% and 33% in early lactation (Kleen et al. 2004; Morgante et al. 2007) and between 18% and 26% in mid-lactation (Kleen et al. 2004; Garret et al. 1997). Table 1 summarizes the prevalence of SARA in dairy herds reported by different authors.
The diagnosis of SARA is difficult under farm conditions as clinical signs are commonly subtle and delayed (Humer et al., 2018). The clinical findings that may direct the veterinarian’s attention to the possible occurrence of SARA have recently been summarized by Oetzel (2017) and include, for example, a poor body condition score and frequent cases of infections. The recommendation is to look at the presence of multiple signs, as the ones listed below, since there is no specific and unique indicator for SARA.
Some symptoms of SARA:
1. Liver abscesses may occur as the result of a cascade of events starting with rumenitis and rumen parakeratosis. Once the rumen epithelium is inflamed, bacteria can leak into portal circulation and causing abscesses. A more specific finding indicative of SARA is liver abscesses at slaughter that may reach prevalences of >30% in cull cows (Rezac et al., 2014). Limit: post mortem information often lost.
2. Variable intake and/or milk production. In cows affected by SARA, a fluctuating feeding pattern has been described as the most consistent symptom. During mid-lactation, variable feed intake may be indicated by the observation of variable milk production; however, during early lactation, this will likely go unnoticed, because of mobilisation of body reserves (Humer et al., 2018).
3. Milk fat depression. The interpretation of low milk fat is quite difficult, as the normal milk fat percentage depends largely on breed, DIM and season. Further, herd means may obscure outlier cows with very low or high milk fat content. Therefore it might be useful to interpret milk fat content as a proportion of cow with very low (<2.5% for Holstein cows) test results; whereby these cows should not represent more than approximately 10% of the herd (Oetzel, 2007). In addition, it is important to keep in mind that also other factors can lead to low milk fat concentration like feeding excessive amount of plant lipids rich in polyunsaturated fatty acids.
4. Alterations in feces and diarrhea. SARA affects the consistency and particle size of feces; however, those alterations are usually transient. Typical feces appearance is bright yellowish with a sweet–sour smell (Kleen et al., 2003). Furthermore, feces may appear foamy with gas bubbles and whole cereal grains as well as higher amounts of undigested fiber might be present. The size of fecal particles may be enlarged being around 1–2 cm instead of the more normal size of less than 0.5 cm (Hall, 2002).
5. High incidence of lameness. During SARA, vasoactive molecules like histamine, LPS and lactic acid, are released into the blood stream. These molecules play an important role in the laminitis etiology, weakening the hoof tissue and predisposing the animals to lameness. Reference values are impossible to set since environmental factors play a big role in such kind of disease.
SARA has long-term devastating health and economic consequences in dairy cattle. The most visible and direct effect of the acidotic stress can be observed in the work published by Khafipour et al. 2009, where a SARA challenge was induced in lactating cows. In this experiment about 20% of a 50:50 forage to concentrate (F:C) ratio TMR, was replaced by a pellet containing 50% barley and 50% wheat for one week, resulting in an F:C of 40:60.
The SARA challenge induced in this experiment reduced the DMI (15%), milk yield (3.3 kg/d), and milk fat (0.12% point).
Few attempts have been done in order to calculate the potential economic impact of SARA. One of the most cited, is a case study conducted on a 500-cow dairy in central New York (Stone, 1999). Stone calculated a cost of $400 to $475 lost income per cow per year due to SARA. This rough estimation was simply calculated multiplying a reduced milk yield by 2.7 kg/day, milk fat by 0.3% point and milk protein by 0.12% points for the entire lactation.
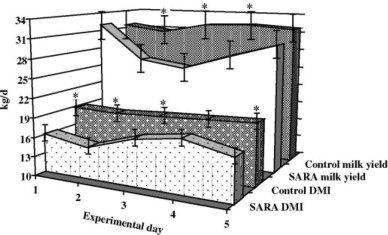
Figure 2. Daily averages of DMI and milk yield in dairy cows fed a basal TMR during control or TMR with wheat-barley pellets during subacute ruminal acidosis (SARA) treatment. Error bars indicate standard error of difference between treatments (SED); within each day, * = P < 0.05.
The performance loss can be explained at physiological level, by the systemic inflammation status occurring in cows subjected to an acidotic challenge. An unbalance between carbohydrate and physically effective fiber causes a shift towards Gram-negative bacteria resulting in the release of cell-free lipopolysaccharide (LPS) in the rumen. LPS can then translocate trough gut and to a lesser extent trough rumen epithelium and reach systemic circulation triggering a strong inflammation response (Zebeli and Metzler-Zebeli, 2012).
However, any estimation on performance loss would be inevitably inaccurate for two main reasons:
Given the lag time following the onset of SARA, there is no specific treatment. This reinforces the importance of prevention.
Besides offering the cows a balanced diet in terms of amount and degradability of carbohydrate and quantity and size of fiber, feeding management is of vital importance to minimize the risk of SARA.
A list of practical management indications have been listed in the review of Humer et al. 2018, and includes:
| Particle fraction | Screen size | TMR 1 (%) | TMR 2 (%) | PMR (%) |
|---|---|---|---|---|
| Large particles | >19 mm | 3-8 | 3-8 | 15-25 |
| Medium particles | 8-19 mm | 30-40 | 35-45 | 35-65 |
| Fine particles | 1.18-8 mm | 30-40 | 40-50 | 15-25 |
| Very fine particles | <1.18 mm | <20 | <10 | <8 |
Table 2. Recommendations for TMR particle size distribution when the TMR is composed of ground concentrates (TMR 1), with pelleted concentrates (TMR 2) or the diet is offered as partial mixed ration (PMR) (partly adapted from Heinrichs and Kononoff, 2002)
Supplementation of feed additives represents another commonly used approach to mitigate the consequences of SARA. Commonly used feed additives include:
One of the most used additive class in this sense are yeasts products which can be provided as live yeasts, dead yeasts, or yeast culture products.
Levabon® Rumen E is an autolyzed yeast, with a prebiotic mode of action, which has shown beneficial effects in cows exposed to an acidotic challenge. In a study, done in cooperation with the University of Veterinary Medicine, Vienna, rumen-cannulated cows were fed a pure forage diet and switched to a 65% concentrate diet on DM basis, to induce an acidotic stress. Supplementing the concentrate rich diet with Levabon® increased duration of eating, total chewing and DMI compared with the control diet (Kröger et al. 2017).
Moreover Levabon® caused pronounced effects on the concentration of biogenic amines, like histamine, during the first acidotic challenge, showing adecrease by 31% compared with control.
Its beneficial effect is also reflected at microbiome level. In the same study Levabon® decreased gram-negative bacteria thanks to the binding activity of certain yeast components like mannan oligosaccharides, β-glucans, chitin, peptides, AA, and nucleotides. The same components also acted as substrate for cellulolytic bacteria, promoting growth of Ruminicoccus and Clostridium spp. in the challenged rumen and contributing to the maintenance of a physiological ruminal pH (Neubauer et al., 2018).
Mycofix® binding complex demonstrated a high affinity towards lipopolysaccharides (LPS) in vitro. LPS binding has been tested both in presence of high aflatoxin concentrations, to exclude competition between Afla and LPS for the same binding sites, and in rumen fluid using Rumen Simulation Technique (RuSiTec). (Image available if needed)
On this basis, in vivo studies have been conducted to test Mycofix® in cows subjected to acidotic stress. In a study done by Prof. Zebeli and his team, non-lactating cows were challenged with intermittent high concentrate diets to induce SARA. As a result of this nutritional stress, levels of liver enzymes such as AST and GLDH, considered markers for hepatocyte integrity of dairy cows (Bobe et al. 2004), increased compared to levels detected in control cows (Figure 3).
The liver plays a major role in the inflammation response caused by LPS release and subsequent translocation in the blood circulation. This mechanism is widely described by Zebeli and Metzler-Zebeli, 2012. The beneficial effect of the clay mineral CM (Mycofix®) supplementation on improved liver health, can be explained by a lower toxic load in the rumen and systemic circulation because of its known capability to absorb LPS (Humer et al., 2019).
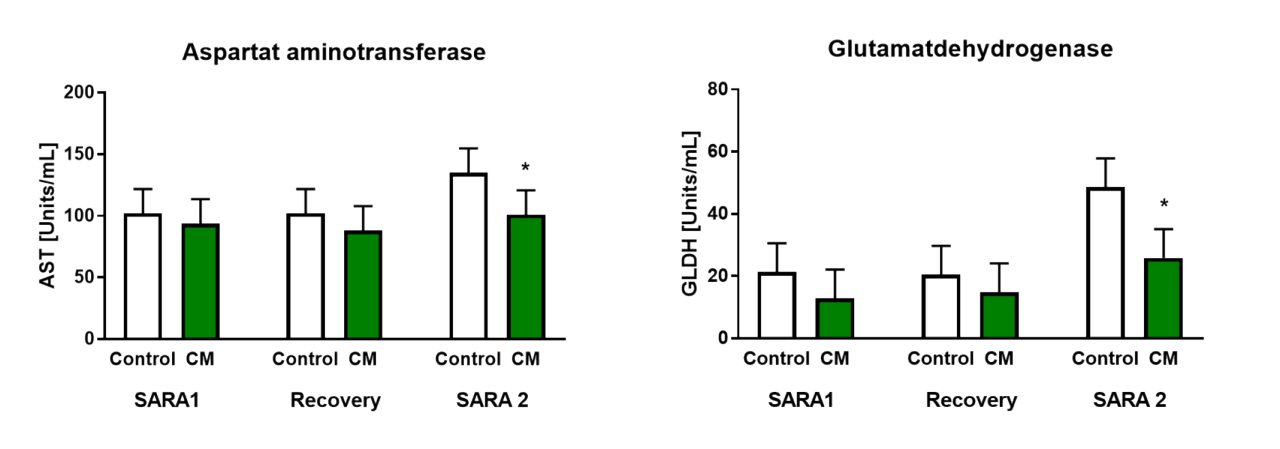
Figure 3. AST and GLDH concentration measured in cows fed control diet and clay mineral additive CM. (Adapted from Humer et al., 2019)
Another interesting effect observed was the significant reduction of certain biogenic amines whose production is increased during high-grain feeding. Among all biogenic amines, histamine, which is also known to play a role in the pathogenesis of laminitis, was significantly reduced by 28% in cows supplemented with Mycofix® (Humer et al., 2019).
Finally, to estimate the global effect of Mycofix® on all the measured parameters, a multivariate analysis was conducted to identify characteristic trends or grouping among cows fed a pure forage diet (Baseline), the SARA diet (65% concentrate; = control) or the diet with Mycofix®. The outcome was that SARA samples clustered separately from the baseline samples, whereby the cows receiving Mycofix® clustered closer to the baseline than control cows (Humer et al., 2019) (Fig. 4).
The beneficial effect of Mycofix® on cows affected by SARA is reflected also at microbiome level. In another paper originating from the same experiment, Mycofix® showed a certain potential to reduce bacteria that are associated with low pH, such as Lactobacillus, and favor high abundant genera such as Campylobacter, Butyrivibrio, and lower abundant Gram-positive commensal bacteria. Along with what has been previously shown in vitro, Mycofix® showed a decreasing effect on possible harmful bacteria, especially Gram-negative genera, such as Treponema, Fusobacteria, and Succiniclasticum. These groups include LPS producing species and potential host pathogens (Neubauer et al. 2019).
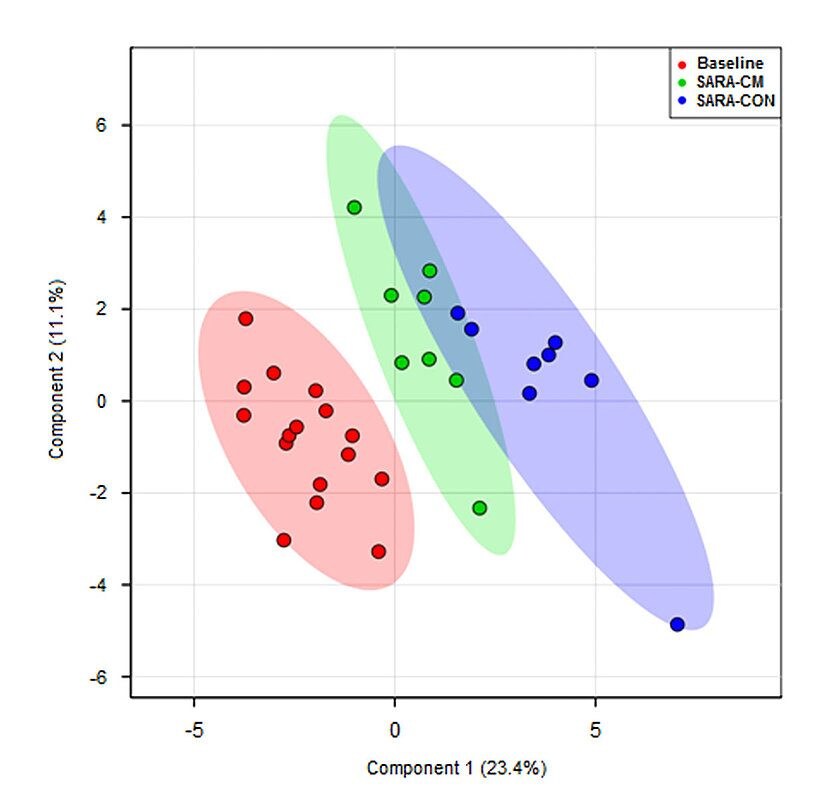
Figure 4. A partial least-squares discriminant analysis (PLS-DA) of the blood metabolites that were affected by the feed additive. The two-dimensional score plot distinguishes the metabolic profiles of cows fed either a pure forage diet (Baseline; red) or a 65% concentrate diet (subacute ruminal acidosis, SARA) without feed additive (control, CON; blue), or a clay mineral-based product (CM; green).
Digestarom® contains a blend of spices, herbs and essential oils that showed potential to modulate reticular pH in different ways. The product was tested in an extensive study done in cooperation with the University of Veterinary Medicine in Vienna, where non-lactating cows were fed an intermittent high concentrate diet to induce SARA.
One of the main finding was that supplemetation of Digestarom® improved reticular pH dynamics when the lowest reticular pH readings were observed in control cows subjected to acidotic stress. In particular Digestarom® increased the time spent ruminating and total chewing (Fig.5) (Kröger et al. 2017).
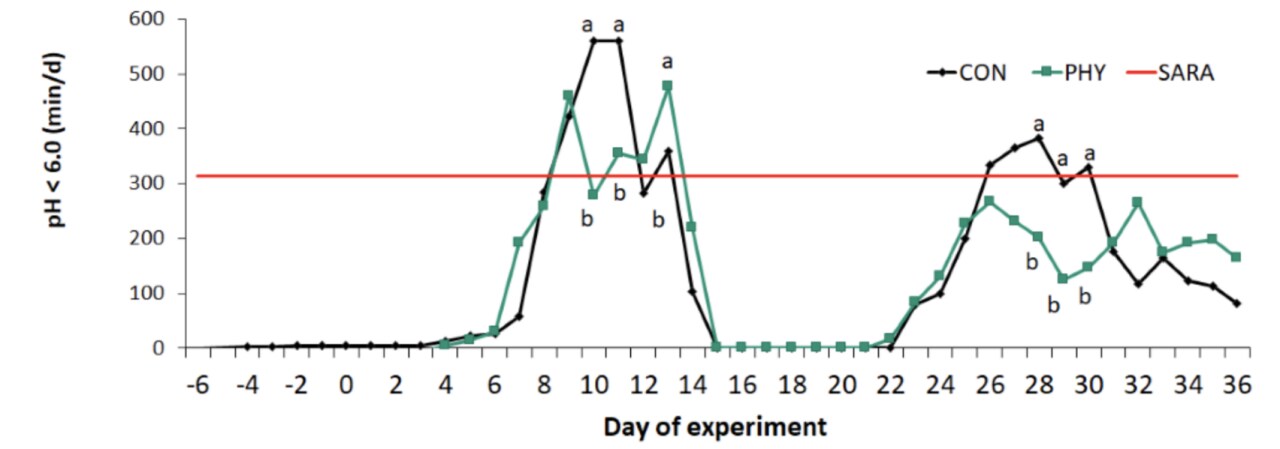
Figure 5. Duration of reticular pH <6.0 in dairy cows fed either a control diet (CON), or a diet supplemented with phytogenic compounds (PHY) per day of experiment. The solid line indicates the SARA threshold of a reticular pH <6.0 for longer than 314 min/d. Treatments with different letters (a,b) differ significantly within the same day (P < 0.05) (Adapted from Kröger et al. 2017).
However, the positive effects of Digestarom® in shortening the time of reticular pH <6.0, cannot be solely attributed to changes in ruminating behavior. In fact, during the second concentrate challenge (CONC 2), there was a positive effect on pH without any influence in chewing variables (Kröger et al. 2017).
The explanation could come from the modulating effect that Digestarom® showed towards ruminal bacteria community with potential effects on reducing the ruminal degradation of starch-rich feeds. Almost all of the bacteria that were decreased by Digestarom® are known starch utilizers, including the genera Shuttleworthia, Olsenella, Bacteroides, Bifidobacterium, Roseburia, and Syntrophococcus (Calsamiglia et al., 2007; Patra, 2011), whereas no fiber fermenting taxa were reduced (Neubauer et al. 2018).
In a high-concentrate diet, the increase in starch provided would normally supply amylolytic bacteria with sufficient substrate to promote growth. The decrease of amylolytic bacteria seen with supplementation of Digestarom® in this experiment, supports the mode of action stated by Calsamiglia et al. (2007) and Cobellis et al. (2016).
A reduction in starch utilizers would potentially delay the onset of SCFA fermentation, reduce SCFA (short-chain fatty acids) accumulation, and thereby prevent rapid and extended duration of low pH after feeding (Neubauer et al. 2018).
Moreover, with a decrease in starch utilizers, cellulolytic bacteria might have a chance to evolve due to less competition (Patra and Yu, 2015). This is supported by the higher reticulo-rumen pH previously reported for Digestarom® (Kröger et al., 2017).
Secrist, D.S., Hill, W.J., Gill, D.R., 1998. Acidosis in cattle: a review. J. Anim. Sci. 76 (1), 275–286., Z.K., 2011. Occurrence of metabolic alkalosis in rumen lactic acidosis: a review article. Comp. Clin. Pathol. 20 (1), 1–3.
Humer E., Aschenbach J.R., Neubauer V., Kröger I., Khiaosa-Ard R, Baumgartner W., Zebeli Q.
Signals for identifying cows at risk of subacute ruminal acidosis in dairy veterinary practice. J Anim Physiol Anim Nutr (Berl). 2018 Apr;102(2):380-392.
Hall, M. B. (2002). Characteristics of manure: What do they mean? In Proceedings of the Tri-State Dairy Nutrition Conference, Fort Wayne, Indiana, US, pp. 141–147.
Oetzel, G. R. (2017). Diagnosis and management of subacute ruminal acidosis in dairy herds. Veterinary Clinics of North America: Food Animal Practice, 33, 463–480.
Oetzel, G.R., 2007. Subacute ruminal acidosis in dairy herds: physiology, pathophysiology, milk fat responses, and nutritional management. Proc. AABP 40th Annual Conference, Vancouver, BC, Canada, pp. 89–119.
Oetzel, G.R., 2003. Subacute ruminal acidosis in dairy cattle. Adv. Dairy Technol. 15, 307–317.
Oetzel, G.R., Nordlund, K.V., Garrett, E.F., 1999. Effect of ruminal pH and stage of lactation on ruminal lactate concentrations in dairy cows. J. Dairy Sci. 82, 38.
Abdela N., 2016. Sub-acute Ruminal Acidosis (SARA) and its Consequence in Dairy Cattle: A Review of Past and Recent Research at Global Prospective. Achievements in the Life Sciences 10 (2016) 187–196
Busquet, P. W. Cardozo, L. Castillejos, and A. Ferret. 2007. Invited review: Essential oils as modifiers of rumen microbial fermentation. J. Dairy Sci. 90:2580–2595.
Bobe G, Young JW and Beitz DC 2004. Invited review: Pathology, etiology, prevention, and treatment of fatty liver in dairy cows. Journal of Dairy Science 87, 3105–3124.
Calsamiglia, S., M. Busquet, P. W. Cardozo, L. Castillejos, and A. Ferret. 2007. Invited review: Essential oils as modifiers of rumen microbial fermentation. J. Dairy Sci. 90:2580–2595. Rehage, J., Noordhuizen,
J.P.T., 2003. Subacute ruminal acidosis (SARA): a review. J. Vet. Med. A Physiol. Pathol. Clin. Med. 50,
Cobellis, G., M. Trabalza-Marinucci, and Z. Yu. 2016. Critical evaluation of essential oils as rumen modifiers in ruminant nutrition: A review. Sci. Total Environ. 545–546:556–568.
Kleen, J.L., Upgang, L., Rehage, J., 2013. Prevalence and consequences of subacute ruminal acidosis in German dairy herds. Acta Vet. Scand. 55, 1–6.
Jing L, Dewanckele L, Vlaeminck B, Van Straalen WM, Koopmans A, Fievez V. Susceptibility of dairy cows to subacute ruminal acidosis is reflected in milk fatty acid proportions, with C18:1 trans-10 as primary and
C15:0 and C18:1 trans-11 as secondary indicators. J Dairy Sci. 2018 Nov;101(11):9827-9840.
Krause, M.K., Otzel, G.R., 2006. Understanding and preventing subacute ruminal acidosis in dairy herds: a review. Anim. Feed Sci. Technol. 126, 215–236.
McCann, J.C., Luan, S., Cardoso, F.C., Derakhshani, H., Khafipour, E., Loor, J.J., 2016. Induction of subacute ruminal acidosis affects the ruminal microbiome and epithelium. Front. Microbiol. 7, 701.
Garrett, E.F., Nordlund, K.V., Goodger,W.J., Oetzel, G.R., 1997. A cross-sectional field study investigating the effect of periparturient dietarymanagement on ruminal pH in early lactation dairy cows. J. Dairy Sci. 80, 169.
O'Grady, L., Doherty, M.L., Mulligan, F.J., 2008. Subacute ruminal acidosis (SARA) in grazing Irish dairy cows. (special issue: production diseases of the transition cow.). Vet. J. 176 (1), 44–49.
Morgante,M., Stelletta, C., Berzaghi, P., Gianesella,M., Andrighetto, I., 2007. Subacute rumen acidosis in lactating cows: an investigation in intensive Italian dairy herds. J. Anim. Physiol. Anim. Nutr. (Berl) 91 (5–6), 226–234.
Neubauer V., Humer E., Mann E., Kröger I., Reisinger N., Wagner M., Zebeli Q., Petri R.M., 2019. Effects clay mineral supplementation on particle-associated and epimural microbiota, and gene expression in the rumen of cows fed high-concentrate diet. Anaerobe 59 (2019) 38-48
Stefańska, B., Nowak,W., Komisarek, J., Taciak,M., Barszcz, M., Skomiał, J., 2016. Prevalence and consequence of subacute ruminal acidosis in polish dairy herds. J. Anim. Physiol. Anim. Nutr.
Devries, T.J., Beauchemin, K.A., Dohme, F., Schwartzkopf Genswein, K.S., 2009. Repeated ruminal acidosis challenges in lactating dairy cows at high and low risk for developing acidosis: feeding, ruminating, and lying behavior. J. Dairy Sci. 92 (10), 5067–5078.
Rezac, D. J., Thomson, D. U., Siemens, M. G., Prouty, F. L., Reinhardt, C. D., & Bartle, S. J. (2014). A survey of gross pathologic conditions in cull cows at slaughter in the Great Lakes region of the United States. Journal of Dairy Science, 97, 4227–35.
Stone, W.C., 2004. Nutritional approaches to minimize subacute ruminal acidosis and laminitis in dairy cattle. J. Dairy Sci. 87, E13–E26.
Enemark, J.M.D., Jorgensen, R.J., Enemark, P.S., 2002. Rumen acidosis with special emphasis on diagnosis aspects of subclinical rumen acidosis: a review. Veterinarija ir Zootechnika 42, 16–29.
Enemark, J.M.D., Jørgensen, R.J., Kristensen, N.B., 2004. An evaluation of parameters for the detection of subclinical rumen acidosis in dairy herds. Vet. Res. Commun. 28 (8), 687–709.
Zebeli Q., Dijkstra J., Tafaj M., Steingass H., Ametaj B. N., and Drochner W., 2008. Modelling the Adequacy of Dietary Fiber in Dairy Cows Based on the Responses of Ruminal pH and Milk Fat Production to Composition of the Diet. J. Dairy Sci. 91:2046–2066
Plaizier JC1, Krause DO, Gozho GN, McBride BW. Subacute ruminal acidosis in dairy cows: the physiological causes, incidence and consequences. Vet J. 2008 Apr;176(1):21-31.
Aceto, H., Simeone, A.J., Fergusson, J.D., 2000. Effect of rumenocentesis on health and productivity in dairy cows. J. Anim. Sci. 78 (Suppl. 1) (Abstr. 162).
Li, S., Gozho, G.N., Gakhar, N., Khafipour, E., Krause, D.O., Plaizier, J.C., 2012. Evaluation of diagnosticmeasures for subacute ruminal acidosis in dairy cows. Can. J. Anim. Sci. 92 (3), 353–364.
Gianesella, M., Massimo, M., Cannizzo, C., Annalisa, S., Paolo, D., Vanessa, M., Elisabetta, G., 2010. Subacute Ruminal Acidosis and Evaluation of Blood Gas Analysis in Dairy Cow, Veterinary Medicine International, 2010.
Khafipour E., Krause D. O., and Plaizier J. C, 2009. A grain-based subacute ruminal acidosis challenge causes translocation of lipopolysaccharide and triggers inflammation. J. Dairy Sci. 92:1060–1070
Li S., Khafipour E., Krause D. O., Kroeker A., Rodriguez-Lecompte J. C., Gozho G. N., Plaizier J. C., 2012. Effects of subacute ruminal acidosis challenges on fermentation and endotoxins in the rumen and hindgut of dairy cows. J. Dairy Sci. 95 :294–303
Zebeli, Q., Metzler-Zebeli, B.U., 2012. Interplay between rumen digestive disorders and diet-induced inflammation in dairy cattle. Res. Vet. Sci. 93 (3), 1099–1108
Kröger, I., E. Humer, V. Neubauer, N. Reisinger, S. Aditya, and Q. Zebeli. 2017. Modulation of chewing behavior and reticular pH in non-lactating cows challenged with concentrate-rich diets supplemented with phytogenic compounds and autolyzed yeast. J. Dairy Sci. 100:9702–9714.
Humer E., Kröger I., Neubauer V., Schedle K., Reisinger N., and Zebeli Q., 2018. Supplementing phytogenic compounds or autolyzed yeast modulates ruminal biogenic amines and plasma metabolome in dry cows experiencing subacute ruminal acidosis. J. Dairy Sci. 101:1–16.
Humer E., Kröger I., Neubauer V., Reisinger N. and Zebeli Q., 2019. Supplementation of a clay mineral-based product modulates plasma metabolomic profile and liver enzymes in cattle fed grain-rich diets. Animal;13(6):1214-1223.
Neubauer V., Petri R., Humer E., Kröger I., Mann E., Reisinger N., Wagner M., and Zebeli Q., 2018. High-grain diets supplemented with phytogenic compounds or autolyzed yeast modulate ruminal bacterial community and fermentation in dry cows. J. Dairy Sci. 101:1–15
Humer E., Petri R. M., Aschenbach J. R., Bradford B. J., Penner G. B., Tafaj M., Südekum K.-H.and Zebeli Q., 2018. Invited review: Practical feeding management recommendations to mitigate the risk of subacute ruminal acidosis in dairy cattle. J. Dairy Sci. 101:1–17.
Biomin® BioStabil is a formulation of strategically selected Lactic Acid Bacteria for optimal haylage, silage and forage preservation. Biomin® BioStabil preserves the energy in your silage.
Levabon® Rumen E is an innovative brewers yeast cattle feed supplement for dairy and beef animals including calves. It delivers an enhanced prebiotic effect compared to conventional yeasts for better productivity.
Our specific blends of essential oil components have shown to help deliver a better balance of gut microflora.
Increase the proportion of ‘good’ bacteria, and you have a better performing gut.
The Biotronic® line of state-of-the-art enhanced acidifiers support modern farm animals against Gram-negative bacteria.
Digestarom® line of phytogenics contain unique blends of herbs and spices, essential oils and other plant extracts to improve palatability and acceptance of feed, support digestion and overall performance of farm animals.
Track feed crop contamination levels anywhere in the world on your device
Our portfolio of tools helps to understand the potential risks of mycotoxins for animal species and location.
The Mycotoxin Prediction Service delivers assessments of expected mycotoxin levels in the upcoming harvest of corn (maize) and wheat around the world.
We offer a range of analytical services to customers to assess the mycotoxin contamination of feed materials.
FUMzyme® Silage is a unique additive sprayed onto corn (maize) at harvest that targets and detoxifies harmful fumonisins, so that the resulting silage is safe and fumonisin-free for livestock nutrition.
The risk management solution against aflatoxins and/or ergot alkaloids in animal feed.
Targeted protection against adsorbable mycotoxins plus fumonisins.
The Mycofix® portfolio of feed additives represents the most state-of-the-art solution for protecting animal health by deactivating mycotoxins that contaminate farm animal feed. Its safety and efficacy are proven by 7 EU authorizations for substances that deactivate mycotoxins.
Absolute protection against the broadest range of mycotoxins.
Absolute protection against the broadest range of mycotoxins. With ZENzyme® Faster and Better
FUMzyme®, the only enzyme that effectively detoxifies fumonisins safely and irreversibly, is available for post-pellet and liquid application in animal feed. From the creators of Mycofix®. Naturally Ahead.
Starting the dsm-firmenich World Mycotoxin Survey already in 2004, we are proud to celebrate a 20 year`s anniversary this year. The increasing sample size over the years gives detailed insights on the incidence of the six major mycotoxins in the agricultural commodities used for livestock feed in order to identify the potential risk posed to livestock animal production.
Only analyzing for single mycotoxins can lead to underestimation of the detrimental effects of mycotoxins on animal health and performance. Our long-term monitoring of mycotoxins in different commodities shows that cooccurrence of mycotoxins is the rule and not the exception. Here we need support of state-of the art analytical methods based on LC-MS/MS. These allow to detect multiple mycotoxins in one run. The high sensitivity of the method is important, as already moderate levels of mycotoxins can have a detrimental effect. This is especially true in case of co-contamination.
Effects of mycotoxins on animals are diverse and range from carcinogenicity, hepatoxicity and neurotoxicity to impaired reproduction, digestive disorders, immunomodulation and decreased performance. Clinical signs can be seen at high levels of mycotoxin contamination but more frequently we observe subclinical effects. Already moderate levels of mycotoxins, especially during chronic exposure, can negatively affect the animals. Mycotoxins influence the immune system, the integrity of the gut barrier and act as predisposing factors for disease.
The dsm-firmenich Mycotoxin Survey constitutes the longest running and most comprehensive data set on mycotoxin occurrence. The survey results provide insights on the incidence of the six major mycotoxins in the agricultural commodities used for livestock feed in order to identify the potential risk posed to livestock animal production.
With our World Mycotoxin Survey results on the current levels of mycotoxin contamination in different commodities and regions of the world, we want to inform and support feed and animal producers to get a better picture of the mycotoxin risk in their region or in imported feed ingredients.




Get in touch with a dsm-firmenich Animal Nutrition & Health specialist or find contacts around the world to suit your needs.
At dsm-firmenich, we love to connect with you.
Follow us on any of the channels below.
